Varolan herşey ussaldır, ya da varolmak için kavramsal olarak belirli olmak gerekir. Usdışı olan varolması olanaksız olandır. Hegel’in “Ussal olan edimseldir, ve edimsel olan ussaldır” sözleri edimselliğe ussal olarak yaklaşmaya çabalayan kafalarda değişik biçimlerde yankılanır. Richard Feynman “Yaratamadığımı anlamam” :: “What I cannot create, I do not understand” diyordu.
Hücrenin anlaşılması hücrenin yaratılması olacaktır.
🛑 HÜCRE
Hücre boyutları 1 ve 100 mikrometre arasında değişir.
Bilinen en küçük hücreler mikoplazma denilen küçük bakteriler grubudur. (çap 0,2 μm; 10−14 gram olan toplam kütlesi 8,000,000,000 hidrojen atomunun kütlesine eşittir; tipik insan hücresi mikoplazmadan 400.000 kat daha büyüktür).
İnsan bedeninde yaklaşık 40 trillion (4×1013) hücre bulunur ve bunlardan yaklaşık 80 milyarı beyne aittir.
Bir mit olarak, hücreler 1665’te Robert Hooke tarafından bulundu.
Sitoplazma hücrede biosentezlere katılan 10.000’den çok değişik molekül türü kapsar.
Hücreler yeryüzünde yaklaşık 3,5 milyar yıl önce ortaya çıktı.
Ökaryotik hücreler çekirdek kapsar.
Prokaryotik hücreler çekirdek ve membran-sınırlı örgeneller kapsamaz.
Ökaryot örgenlikler tek- ya da çok-hücreli olabilir.
Prokaryotlar tek-hücrelidir.
Prokaryotlar bakteri ve arkheaları (üç yaşam alanından ikisi) kapsar (?).
Prokaryotlar hücre imleşimi kapsar.
Prokaryotlarda hücre kılıfı hücre membran ve duvarından oluşur (duvar bir kapsül ile kuşatılı da olabilir).
Prokaryotlarda genom (DNA) sitoplazmada bulunur.
Ökaryotlarda bölümlü yapılanma vardır (membran-sınırlı örgeneller).
📹📹📹 CELL ORGANELLES (VİDEO)
📹 The Secret Life of a Cell, Part 1 — Organelles (VİDEO)
📹 The Secret Life of a Cell, Part 1 — Organelles (LINK)
Each cell in the human body is specialized to have a specific job and works together with other cells to orchestrate the symphony of unconscious biological processes. Part 1 of this new series introduces the cell membrane, cytoplasm, and the organelles such as the cytoskeleton, microtubules, centrosome, and vesicles, the basic structures of animal cells, and describes their general function.
📹 The Secret Life of a Cell, Part 2 — Organelles (cont'd) (VİDEO)
📹 The Secret Life of a Cell, Part 2 — Organelles (cont'd) (LINK)
Part 2 of this series continues describing the organelles, the basic tiny structures of eukaryotic cells: lysosomes (which contain over 50 different enzymes), endoplasmic reticulum, ribosomes, Golgi apparatus, and mitochondria (the main energy source for the majority of cellular functions).
📹 The Secret Life of a Cell, Part 3 — The Nucleus (VİDEO)
📹 The Secret Life of a Cell, Part 3 / The Nucleus — (LINK)
Part 3 of the Secret Life of a Cell series introduces the largest structure inside a eukaryotic animal cell, the nucleus. Protected by a plasma membrane, the nucleus houses another structure called the nucleolus, home to the vast majority of genes and the primary site of ribosome synthesis. The nucleus is a densely packed conglomerate of chromatin regulatory proteins called histones, which organize strands of DNA into chromosomes and autosomes. Part 3 also discusses the structure of a chromosome, keeper of the cell's genetic code.
📹📹📹 CELL 2 (VİDEO)
📹 l Cell structure and Function — Animal cell and Plant cell / creative Learning (VİDEO)
📹 Cell structure and Function — Animal cell and Plant cell / creative Learning (LINK)
Cell Structure and Function
The cell membrane performs many important functions within the cell such as osmosis, diffusion, transport of nutrients into the cell, processes of ingestion and secretion. The cell membrane is strong enough to provide the cell with mechanical support and flexible enough to allow cells to grow and move.
Explore some examples of specialized plant and animal cells with the Amoeba Sisters! Video explains how specialized cell structure suits their function. Expand details for table of contents.
📹 Prokaryotic vs. Eukaryotic Cells (Updated) / Amobea Sisters (VİDEO)
📹 Prokaryotic vs. Eukaryotic Cells (Updated) / Amobea Sisters (LINK)
This Amoeba Sisters video starts with providing examples of prokaryotes and eukaryotes before comparing and contrasting prokaryotic cells with eukaryotic cells!
Principal structures of an animal cellCytoplasm surrounds the cell's specialized structures, or organelles. Ribosomes, the sites of protein synthesis, are found free in the cytoplasm or attached to the endoplasmic reticulum, through which materials are transported throughout the cell. Energy needed by the cell is released by the mitochondria. The Golgi complex, stacks of flattened sacs, processes and packages materials to be released from the cell in secretory vesicles. Digestive enzymes are contained in lysosomes. Peroxisomes contain enzymes that detoxify dangerous substances. The centrosome contains the centrioles, which play a role in cell division. The microvilli are fingerlike extensions found on certain cells. Cilia, hairlike structures that extend from the surface of many cells, can create movement of surrounding fluid. The nuclear envelope, a double membrane surrounding the nucleus, contains pores that control the movement of substances into and out of the nucleoplasm. Chromatin, a combination of DNA and proteins that coil into chromosomes, makes up much of the nucleoplasm. The dense nucleolus is the site of ribosome production.
Cell, in biology, the basic membrane-bound unit that contains the fundamental molecules of life and of which all living things are composed. A single cell is often a complete organism in itself, such as a bacterium or yeast. Other cells acquire specialized functions as they mature. These cells cooperate with other specialized cells and become the building blocks of large multicellular organisms, such as humans and other animals. Although cells are much larger than atoms, they are still very small. The smallest known cells are a group of tiny bacteria called mycoplasmas; some of these single-celled organisms are spheres as small as 0.2 μm in diameter (1μm = about 0.000039 inch), with a total mass of 10−14 gram—equal to that of 8,000,000,000 hydrogen atoms. Cells of humans typically have a mass 400,000 times larger than the mass of a single mycoplasma bacterium, but even human cells are only about 20 μm across. It would require a sheet of about 10,000 human cells to cover the head of a pin, and each human organism is composed of more than 30,000,000,000,000 cells.
This article discusses the cell both as an individual unit and as a contributing part of a larger organism. As an individual unit, the cell is capable of metabolizing its own nutrients, synthesizing many types of molecules, providing its own energy, and replicating itself in order to produce succeeding generations. It can be viewed as an enclosed vessel, within which innumerable chemical reactions take place simultaneously. These reactions are under very precise control so that they contribute to the life and procreation of the cell. In a multicellular organism, cells become specialized to perform different functions through the process of differentiation. In order to do this, each cell keeps in constant communication with its neighbours. As it receives nutrients from and expels wastes into its surroundings, it adheres to and cooperates with other cells. Cooperative assemblies of similar cells form tissues, and a cooperation between tissues in turn forms organs, which carry out the functions necessary to sustain the life of an organism.
Special emphasis is given in this article to animal cells, with some discussion of the energy-synthesizing processes and extracellular components peculiar to plants. (For detailed discussion of the biochemistry of plant cells, seephotosynthesis. For a full treatment of the genetic events in the cell nucleus, seeheredity.)
Animal cells and plant cells contain membrane-bound organelles, including a distinct nucleus. In contrast, bacterial cells do not contain organelles.
.
A cell is enclosed by a plasma membrane, which forms a selective barrier that allows nutrients to enter and waste products to leave. The interior of the cell is organized into many specialized compartments, or organelles, each surrounded by a separate membrane. One major organelle, the nucleus, contains the genetic information necessary for cell growth and reproduction. Each cell contains only one nucleus, whereas other types of organelles are present in multiple copies in the cellular contents, or cytoplasm. Organelles include mitochondria, which are responsible for the energy transactions necessary for cell survival; lysosomes, which digest unwanted materials within the cell; and the endoplasmic reticulum and the Golgi apparatus, which play important roles in the internal organization of the cell by synthesizing selected molecules and then processing, sorting, and directing them to their proper locations. In addition, plant cells contain chloroplasts, which are responsible for photosynthesis, whereby the energy of sunlight is used to convert molecules of carbon dioxide (CO2) and water (H2O) into carbohydrates. Between all these organelles is the space in the cytoplasm called thecytosol. The cytosol contains an organized framework of fibrous molecules that constitute the cytoskeleton,which gives a cell its shape, enables organelles to move within the cell, and provides a mechanism by which the cell itself can move. The cytosol also contains more than 10,000 different kinds of molecules that are involved in cellular biosynthesis, the process of making large biological molecules from small ones.
Eukaryotic cell.
Cutaway drawing of a eukaryotic cell.
Specialized organelles are a characteristic of cells of organisms known as eukaryotes. In contrast, cells of organisms known as prokaryotes do not contain organelles and are generally smaller than eukaryotic cells. However, all cells share strong similarities in biochemical function.
Cells contain a special collection of molecules that are enclosed by a membrane. These molecules give cells the ability to grow and reproduce. The overall process of cellular reproduction occurs in two steps: cell growth and cell division. During cell growth, the cell ingests certain molecules from its surroundings by selectively carrying them through its cell membrane. Once inside the cell, these molecules are subjected to the action of highly specialized, large, elaborately folded molecules called enzymes.Enzymes act as catalysts by binding to ingested molecules and regulating the rate at which they are chemically altered. These chemical alterations make the molecules more useful to the cell. Unlike the ingested molecules, catalysts are not chemically altered themselves during the reaction, allowing one catalyst to regulate a specific chemical reaction in many molecules.
Biological catalysts create chains of reactions. In other words, a molecule chemically transformed by one catalyst serves as the starting material, or substrate, of a second catalyst and so on. In this way, catalysts use the small molecules brought into the cell from the outside environment to create increasingly complex reaction products. These products are used for cell growth and the replication of genetic material. Once the genetic material has been copied and there are sufficient molecules to support cell division, the cell divides to create two daughter cells. Through many such cycles of cell growth and division, each parent cell can give rise to millions of daughter cells, in the process converting large amounts of inanimate matter into biologically active molecules.
Cells are largely composed of compounds that contain carbon. The study of how carbon atoms interact with other atoms in molecular compounds forms the basis of the field of organic chemistry and plays a large role in understanding the basic functions of cells. Because carbon atoms can form stable bonds with four other atoms, they are uniquely suited for the construction of complex molecules. These complex molecules are typically made up of chains and rings that contain hydrogen,oxygen, and nitrogen atoms, as well as carbon atoms. These molecules may consist of anywhere from 10 to millions of atoms linked together in specific arrays. Most, but not all, of the carbon-containing molecules in cells are built up from members of one of four different families of small organic molecules: sugars, amino acids,nucleotides, and fatty acids. Each of these families contains a group of molecules that resemble one another in both structure and function. In addition to other important functions, these molecules are used to build large macromolecules. For example, the sugars can be linked to form polysaccharides such as starch and glycogen, the amino acids can be linked to form proteins, the nucleotides can be linked to form the DNA (deoxyribonucleic acid) and RNA (ribonucleic acid) of chromosomes, and the fatty acids can be linked to form the lipids of all cell membranes.
Approximate chemical composition of a typical mammalian cell
Aside from water, which forms 70 percent of a cell’s mass, a cell is composed mostly of macromolecules. By far the largest portion of macromolecules are the proteins. An average-sized proteinmacromolecule contains a string of about 400 amino acid molecules. Each amino acid has a different side chain of atoms that interact with the atoms of side chains of other amino acids. These interactions are very specific and cause the entire protein molecule to fold into a compact globular form. In theory, nearly an infinite variety of proteins can be formed, each with a different sequence of amino acids. However, nearly all these proteins would fail to fold in the unique ways required to form efficient functional surfaces and would therefore be useless to the cell. The proteins present in cells of modern animals and humans are products of a long evolutionary history, during which the ancestor proteins were naturally selected for their ability to fold into specific three-dimensional forms with unique functional surfaces useful for cell survival.
Most of the catalytic macromolecules in cells are enzymes. The majority of enzymes are proteins. Key to the catalytic property of an enzyme is its tendency to undergo a change in its shape when it binds to its substrate, thus bringing together reactive groups on substrate molecules. Some enzymes are macromolecules of RNA, called ribozymes. Ribozymes consist of linear chains of nucleotides that fold in specific ways to form unique surfaces, similar to the ways in which proteins fold. As with proteins, the specific sequence of nucleotide subunits in an RNA chain gives each macromolecule a unique character. RNA molecules are much less frequently used as catalysts in cells than are protein molecules, presumably because proteins, with the greater variety of amino acid side chains, are more diverse and capable of complex shape changes. However, RNA molecules are thought to have preceded protein molecules during evolution and to have catalyzed most of the chemical reactions required before cells could evolve (see belowThe evolution of cells).
Cells must obey the laws of chemistry and thermodynamics. When two molecules react with each other inside a cell, their atoms are rearranged, forming different molecules as reaction products and releasing or consuming energy in the process. Overall, chemical reactions occur only in one direction; that is, the final reaction product molecules cannot spontaneously react, in a reversal of the original process, to reform the original molecules. This directionality of chemical reactions is explained by the fact that molecules only change from states of higher free energy to states of lower free energy. Free energy is the ability to perform work (in this case, the “work” is the rearrangement of atoms in the chemical reaction). When work is performed, some free energy is used and lost, with the result that the process ends at lower free energy. To use a familiar mechanical analogy,water at the top of a hill has the ability to perform the “work” of flowing downhill (i.e., it has high free energy), but, once it has flowed downhill, it cannot flow back up (i.e., it is in a state of low free energy). However, through another work process—that of a pump, for example—the water can be returned to the top of the hill, thereby recovering its ability to flow downhill. In thermodynamic terms, the free energy of the water has been increased by energy from an outside source (i.e., the pump). In the same way, the product molecules of a chemical reaction in a cell cannot reverse the reaction and return to their original state unless energy is supplied by coupling the process to another chemical reaction.
All catalysts, including enzymes, accelerate chemical reactions without affecting their direction. To return to the mechanical analogy, enzymes cannot make water flow uphill, although they can provide specific pathways for a downhill flow. Yet most of the chemical reactions that the cell needs to synthesize new molecules necessary for its growth require an uphill flow. In other words, the reactions require more energy than their starting molecules can provide.
Cells use a single strategy over and over again in order to get around the limitations of chemistry: they use the energy from an energy-releasing chemical reaction to drive an energy-absorbing reaction that would otherwise not occur. A useful mechanical analogy might be a mill wheel driven by the water in a stream. The water, in order to flow downhill, is forced to flow past the blades of the wheel, causing the wheel to turn. In this way, part of the energy from the moving stream is harnessed to move a mill wheel, which may be linked to a winch. As the winch turns, it can be used to pull a heavy load uphill. Thus, the energy-absorbing (but useful) uphill movement of a load can be driven by coupling it directly to the energy-releasing flow of water.
In cells, enzymes play the role of mill wheels by coupling energy-releasing reactions with energy-absorbing reactions. As discussed below, in cells the most important energy-releasing reaction serving a role similar to that of the flowing stream is the hydrolysis of adenosine triphosphate (ATP). In turn, the production of ATP molecules in the cells is an energy-absorbing reaction that is driven by being coupled to the energy-releasing breakdown of sugar molecules. In retracing this chain of reactions, it is necessary first to understand the source of the sugar molecules.
Sugar molecules are produced by the process of photosynthesis in plants and certain bacteria. These organisms lie at the base of the food chain, in that animals and other nonphotosynthesizing organisms depend on them for a constant supply of life-supporting organic molecules. Humans, for example, obtain these molecules by eating plants or other organisms that have previously eaten food derived from photosynthesizing organisms.
Plants and photosynthetic bacteria are unique in their ability to convert the freely available electromagnetic energy in sunlight into chemical bond energy, the energy that holds atoms together in molecules and is transferred or released in chemical reactions. The process of photosynthesis can be summarized by the following equation:
(solar) energy + CO2 + H2O → sugar molecules + O2.
The energy-absorbing photosynthetic reaction is the reverse of the energy-releasing oxidative decomposition of sugar molecules. During photosynthesis, chlorophyll molecules absorb energy from sunlight and use it to fuel the production of simple sugars and other carbohydrates. The resulting abundance of sugar molecules and related biological products makes possible the existence of nonphotosynthesizing life on Earth.
Certain enzymes catalyze the breakdown of organic foodstuffs. Once sugars are transported into cells, they either serve as building blocks in the form of amino acids for proteins and fatty acids for lipids or are subjected to metabolic pathways to provide the cell with ATP. ATP, the common carrier of energy inside the cell, is made from adenosine diphosphate (ADP) and inorganic phosphate (Pi). Stored in the chemical bond holding the terminal phosphate compound onto the ATP molecule is the energy derived from the breakdown of sugars. The removal of the terminal phosphate, through the water-mediated reaction called hydrolysis, releases this energy, which in turn fuels a large number of crucial energy-absorbing reactions in the cell. Hydrolysis can be summarized as follows:
ATP +H2O → ADP + Pi+ energy.
The formation of ATP is the reverse of this equation, requiring the addition of energy. The central cellular pathway of ATP synthesis begins with glycolysis, a form of fermentation in which the sugar glucose is transformed into other sugars in a series of nine enzymatic reactions, each successive reaction involving an intermediate sugar containing phosphate. In the process, the six-carbon glucose is converted into two molecules of the three-carbon pyruvic acid. Some of the energy released through glycolysis of each glucose molecule is captured in the formation of two ATP molecules.
The second stage in the metabolism of sugars is a set of interrelated reactions called the tricarboxylic acid cycle. This cycle takes the three-carbon pyruvic acid produced in glycolysis and uses its carbon atoms to form carbon dioxide (CO2) while transferring its hydrogen atoms to special carrier molecules, where they are held in high-energy linkage.
Tricarboxylic acid cycle.
The eight-step tricarboxylic acid cycle.
In the third and last stage in the breakdown of sugars, oxidative phosphorylation, the high-energy hydrogen atoms are first separated into protons and high-energy electrons. The electrons are then passed from one electron carrier to another by means of an electron-transport chain. Each electron carrier in the chain has an increasing affinity for electrons, with the final electron acceptor being molecular oxygen (O2). As separated electrons and protons, the hydrogen atoms are transferred to O2 to form water. This reaction releases a large amount of energy, which drives the synthesis of a large number of ATP molecules from ADP and Pi. (For further discussion of the electron-transport chain, see belowMetabolic functions.)
Most of the cell’s ATP is produced when the products of glycolysis are oxidized completely by a combination of the tricarboxylic acid cycle and oxidative phosphorylation. The process of glycolysis alone produces relatively small amounts of ATP. Glycolysis is an anaerobic reaction; that is, it can occur even in the absence of oxygen. The tricarboxylic acid cycle and oxidative phosphorylation, on the other hand, require oxygen. Glycolysis forms the basis of anaerobic fermentation, and it presumably was a major source of ATP for early life on Earth, when very little oxygen was available in the atmosphere. Eventually, however, bacteria evolved that were able to carry out photosynthesis. Photosynthesis liberated these bacteria from a dependence on the metabolism of organic materials that had accumulated from natural processes, and it also released oxygen into the atmosphere. Over a prolonged period of time, the concentration of molecular oxygen increased until it became freely available in the atmosphere. The aerobic tricarboxylic acid cycle and oxidative phosphorylation then evolved, and the resulting aerobic cells made much more efficient use of foodstuffs than their anaerobic ancestors, because they could convert much larger amounts of chemical bond energy into ATP.
Cells can thus be seen as a self-replicating network of catalytic macromolecules engaged in a carefully balanced series of energy conversions that drive biosynthesis and cell movement. But energy alone is not enough to make self-reproduction possible; the cell must contain detailed instructions that dictate exactly how that energy is to be used. These instructions are analogous to the blueprints that a builder uses to construct a house; in the case of cells, however, the blueprints themselves must be duplicated along with the cell before it divides, so that each daughter cell can retain the instructions that it needs for its own replication. These instructions constitute the cell’s heredity.
During the early 19th century, it became widely accepted that all living organisms are composed of cells arising only from the growth and division of other cells. The improvement of the microscope then led to an era during which many biologists made intensive observations of the microscopic structure of cells. By 1885 a substantial amount of indirect evidence indicated that chromosomes—dark-staining threads in the cell nucleus—carried the information for cell heredity. It was later shown that chromosomes are about half DNA and half protein by weight.
DNA structure
The initial proposal of the structure of DNA by James Watson and Francis Crick was accompanied by a suggestion on the means of replication.
The revolutionary discovery suggesting that DNA molecules could provide the information for their own replication came in 1953, when American geneticist and biophysicist James Watson and British biophysicist Francis Crick proposed a model for the structure of the double-stranded DNA molecule (called the DNA double helix). In this model, each strand serves as a template in the synthesis of a complementary strand. Subsequent research confirmed the Watson and Crick model of DNA replication and showed that DNA carries the genetic information for reproduction of the entire cell.
All of the genetic information in a cell was initially thought to be confined to the DNA in the chromosomes of the cell nucleus. Later discoveries identified small amounts of additional genetic information present in the DNA of much smaller chromosomes located in two types of organelles in the cytoplasm. These organelles are the mitochondria in animal cells and the mitochondria and chloroplasts in plant cells. The special chromosomes carry the information coding for a few of the many proteins and RNA molecules needed by the organelles. They also hint at the evolutionary origin of these organelles, which are thought to have originated as free-living bacteria that were taken up by other organisms in the process of symbiosis.
It is possible for RNA to replicate itself by mechanisms related to those used by DNA, even though it has a single-stranded instead of a double-stranded structure. In early cells RNA is thought to have replicated itself in this way. However, all of the RNA in present-day cells is synthesized by special enzymes that construct a single-stranded RNA chain by using one strand of the DNA helix as a template. Although RNA molecules are synthesized in the cell nucleus, where the DNA is located, most of them are transported to the cytoplasm before they carry out their functions.
Messenger RNA; translation.
Molecular genetics emerged from the realization that DNA and RNA constitute the genetic material of all living organisms. (1) DNA, located in the cell nucleus, is made up of nucleotides that contain the bases adenine (A), thymine (T), guanine (G), and cytosine (C). (2) RNA, which contains uracil (U) instead of thymine, transports the genetic code to protein-synthesizing sites in the cell. (3) Messenger RNA (mRNA) then carries the genetic information to ribosomes in the cell cytoplasm that translate the genetic information into molecules of protein.
The RNA molecules in cells have two main roles. Some, the ribozymes, fold up in ways that allow them to serve as catalysts for specific chemical reactions. Others serve as “messenger RNA,” which provides templates specifying the synthesis of proteins. Ribosomes, tiny protein-synthesizing machines located in the cytoplasm, “read” the messenger RNA molecules and “translate” them into proteins by using the genetic code. In this translation, the sequence of nucleotides in the messenger RNA chain is decoded three nucleotides at a time, and each nucleotide triplet (called a codon) specifies a particular amino acid. Thus, a nucleotide sequence in the DNA specifies a protein provided that a messenger RNA molecule is produced from that DNA sequence. Each region of the DNA sequence specifying a protein in this way is called a gene.
By the above mechanisms, DNA molecules catalyze not only their own duplication but also dictate the structures of all protein molecules. A single human cell contains about 10,000 different proteins produced by the expression of 10,000 different genes. Actually, a set of human chromosomes is thought to contain DNA with enough information to express between 30,000 and 100,000 proteins, but most of these proteins seem to be made only in specialized types of cells and are therefore not present throughout the body. (For further discussion, see belowThe nucleus.)
A cell with its many different DNA, RNA, and protein molecules is quite different from a test tube containing the same components. When a cell is dissolved in a test tube, thousands of different types of molecules randomly mix together. In the living cell, however, these components are kept in specific places, reflecting the high degree of organization essential for the growth and division of the cell. Maintaining this internal organization requires a continuous input of energy, because spontaneous chemical reactions always create disorganization. Thus, much of the energy released by ATP hydrolysis fuels processes that organize macromolecules inside the cell.
When a eukaryotic cell is examined at high magnification in an electron microscope, it becomes apparent that specific membrane-bound organelles divide the interior into a variety of subcompartments. Although not detectable in the electron microscope, it is clear from biochemical assays that each organelle contains a different set of macromolecules. This biochemical segregation reflects the functional specialization of each compartment. Thus, the mitochondria, which produce most of the cell’s ATP, contain all of the enzymes needed to carry out the tricarboxylic acid cycle and oxidative phosphorylation. Similarly, the degradative enzymes needed for the intracellular digestion of unwanted macromolecules are confined to the lysosomes.
The relative volumes occupied by some cellular compartments in a typical liver cell
cellular compartment
percent of total cell volume
approximate number per cell
cytosol
54
1
mitochondrion
22
1,700
endoplasmic reticulum plus Golgi apparatus
15
1
nucleus
6
1
lysosome
1
300
It is clear from this functional segregation that the many different proteins specified by the genes in the cell nucleus must be transported to the compartment where they will be used. Not surprisingly, the cell contains an extensive membrane-bound system devoted to maintaining just this intracellular order. The system serves as a post office, guaranteeing the proper routing of newly synthesized macromolecules to their proper destinations.
All proteins are synthesized on ribosomes located in the cytosol. As soon as the first portion of the amino acid sequence of a protein emerges from the ribosome, it is inspected for the presence of a short “endoplasmic reticulum (ER) signal sequence.” Those ribosomes making proteins with such a sequence are transported to the surface of the ER membrane, where they complete their synthesis; the proteins made on these ribosomes are immediately transferred through the ER membrane to the inside of the ER compartment. Proteins lacking the ER signal sequence remain in the cytosol and are released from the ribosomes when their synthesis is completed. This chemical decision process places some newly completed protein chains in the cytosol and others within an extensive membrane-bounded compartment in the cytoplasm, representing the first step in intracellular protein sorting.
The newly made proteins in both cell compartments are then sorted further according to additional signal sequences that they contain. Some of the proteins in the cytosol remain there, while others go to the surface of mitochondria or (in plant cells) chloroplasts, where they are transferred through the membranes into the organelles. Subsignals on each of these proteins then designate exactly where in the organelle the protein belongs. The proteins initially sorted into the ER have an even wider range of destinations. Some of them remain in the ER, where they function as part of the organelle. Most enter transport vesicles and pass to the Golgi apparatus, separate membrane-bounded organelles that contain at least three subcompartments. Some of the proteins are retained in the subcompartments of the Golgi, where they are utilized for functions peculiar to that organelle. Most eventually enter vesicles that leave the Golgi for other cellular destinations such as the cell membrane, lysosomes, or special secretory vesicles. (For further discussion, see belowInternal membranes.)
Formation of a multicellular organism starts with a small collection of similar cells in an embryo and proceeds by continuous cell division and specialization to produce an entire community of cooperating cells, each with its own role in the life of the organism. Through cell cooperation, the organism becomes much more than the sum of its component parts.
Early stages of human development.
The ovum contains a small collection of cells in the early stages of human development. As cells divide (A–D), they are separated into different regions of the ovum. Each region of the ovum transmits a unique set of chemical signals to nearby cells. Thus, the signals detected by one cell differ from those detected by its neighbour cells. In this process, known as cell determination, cells are individually programmed to direct them toward development into different cell types.
A fertilized egg multiplies and produces a whole family of daughter cells, each of which adopts a structure and function according to its position in the entire assembly. All of the daughter cells contain the same chromosomes and therefore the same genetic information. Despite this common inheritance, different types of cells behave differently and have different structures. In order for this to be the case, they must express different sets of genes, so that they produce different proteins despite their identical embryological ancestors.
During the development of an embryo, it is not sufficient for all the cell types found in the fully developed individual simply to be created. Each cell type must form in the right place at the right time and in the correct proportion; otherwise, there would be a jumble of randomly assorted cells in no way resembling an organism. The orderly development of an organism depends on a process called cell determination, in which initially identical cells become committed to different pathways of development. A fundamental part of cell determination is the ability of cells to detect different chemicals within different regions of the embryo. The chemical signals detected by one cell may be different from the signals detected by its neighbour cells. The signals that a cell detects activate a set of genes that tell the cell to differentiate in ways appropriate for its position within the embryo. The set of genes activated in one cell differs from the set of genes activated in the cells around it. The process of cell determination requires an elaborate system of cell-to-cell communication in early embryos.
Intrinsic proteins penetrate and bind tightly to the lipid bilayer, which is made up largely of phospholipids and cholesterol and which typically is between 4 and 10 nanometers (nm; 1 nm = 10−9 metre) in thickness. Extrinsic proteins are loosely bound to the hydrophilic (polar) surfaces, which face the watery medium both inside and outside the cell. Some intrinsic proteins present sugar side chains on the cell's outer surface.
A thin membrane, typically between 4 and 10 nanometers (nm; 1 nm = 10−9 metre) in thickness, surrounds every living cell, delimiting the cell from the environment around it. Enclosed by this cell membrane (also known as the plasma membrane) are the cell’s constituents, often large, water-soluble, highly charged molecules such as proteins,nucleic acids,carbohydrates, and substances involved in cellular metabolism. Outside the cell, in the surrounding water-based environment, are ions,acids, and alkalis that are toxic to the cell, as well as nutrients that the cell must absorb in order to live and grow. The cell membrane, therefore, has two functions: first, to be a barrier keeping the constituents of the cell in and unwanted substances out and, second, to be a gate allowing transport into the cell of essential nutrients and movement from the cell of waste products.
Most current knowledge about the biochemical constituents of cell membranes originates in studies of red blood cells. The chief advantage of these cells for experimental purposes is that they may be obtained easily in large amounts and that they have no internal membranous organelles to interfere with study of their cell membranes. Careful studies of these and other cell types have shown that all membranes are composed of proteins and fatty-acid-based lipids. Membranes actively involved in metabolism contain a higher proportion of protein; thus, the membrane of the mitochondrion, the most rapidly metabolizing organelle of the cell, contains as much as 75 percent protein, while the membrane of the Schwann cell, which forms an insulating sheath around many nerve cells, has as little as 20 percent protein.
General structural formula of a glycerophospholipid. The composition of the specific molecule depends on the chemical group (designated R3 in the diagram) linked to the phosphate and glycerol “head” and also on the lengths of the fatty acid “tails” (R1 and R2).
Liposome.
Phospholipids can be used to form artificial structures called liposomes, which are double-walled hollow spheres useful for encapsulating other molecules such as pharmaceutical drugs.
Lipid bilayer; cell membrane.
Phospholipid molecules, like molecules of many lipids, are composed of a hydrophilic “head” and one or more hydrophobic “tails.” In a water medium, the molecules form a lipid bilayer, or two-layered sheet, in which the heads are turned toward the watery medium and the tails are sheltered inside, away from the water. This bilayer is the basis of the membranes of living cells.
Membrane lipids are principally of two types, phospholipids and sterols (generally cholesterol). Both types share the defining characteristic of lipids—they dissolve readily in organic solvents—but in addition they both have a region that is attracted to and soluble in water. This “amphiphilic” property (having a dual attraction; i.e., containing both a lipid-soluble and a water-soluble region) is basic to the role of lipids as building blocks of cellular membranes. Phospholipid molecules have a head (often of glycerol) to which are attached two long fatty acid chains that look much like tails. These tails are repelled by water and dissolve readily in organic solvents, giving the molecule its lipid character. To another part of the head is attached a phosphoryl group with a negative electrical charge; to this group in turn is attached another group with a positive or neutral charge. This portion of the phospholipid dissolves in water, thereby completing the molecule’s amphiphilic character. In contrast, sterols have a complex hydrocarbon ring structure as the lipid-soluble region and a hydroxyl grouping as the water-soluble region.
When dry phospholipids, or a mixture of such phospholipids and cholesterol, are immersed in water under laboratory conditions, they spontaneously form globular structures called liposomes. Investigation of the liposomes shows them to be made of concentric spheres, one sphere inside of another and each forming half of a bilayered wall. A bilayer is composed of two sheets of phospholipid molecules with all of the molecules of each sheet aligned in the same direction. In a water medium, the phospholipids of the two sheets align so that their water-repellent, lipid-soluble tails are turned and loosely bonded to the tails of the molecules on the other sheet. The water-soluble heads turn outward into the water, to which they are chemically attracted. In this way, the two sheets form a fluid, sandwichlike structure, with the fatty acid chains in the middle mingling in an organic medium while sealing out the water medium.
This type of lipid bilayer, formed by the self-assembly of lipid molecules, is the basic structure of the cell membrane. It is the most stable thermodynamic structure that a phospholipid-water mixture can take up: the fatty acid portion of each molecule dissolved in the organic phase formed by the identical regions of the other molecules and the water-attractive regions surrounded by water and facing away from the fatty acid regions. The chemical affinity of each region of the amphiphilic molecule is thus satisfied in the bilayer structure.
Membrane proteins are also of two general types. One type, called the extrinsic proteins, is loosely attached by ionic bonds or calcium bridges to the electrically charged phosphoryl surface of the bilayer. They can also attach to the second type of protein, called the intrinsic proteins. The intrinsic proteins, as their name implies, are firmly embedded within the phospholipid bilayer. Almost all intrinsic proteins contain special amino acid sequences, generally about 20- to 24-amino acids long, that extend through the internal regions of the cell membrane.
Most intrinsic and extrinsic proteins bear on their outer surfaces side chains of complex sugars, which extend into the aqueous environment around the cell. For this reason, these proteins are often referred to as glycoproteins. Some glycoproteins are involved in cell-to-cell recognition (see belowThe cell matrix and cell-to-cell communication).
A scanning electron micrograph of a pancreatic acinar cell, showing mitochondria (blue), rough endoplasmic reticulum (yellow; ribosomes appear as small dots), and Golgi apparatus (gray, at centre and lower left).
One of the triumphs of cell biology during the decade from 1965 to 1975 was the recognition of the cell membrane as a fluid collection of amphiphilic molecules. This array of proteins, sterols, and phospholipids is organized into a liquid crystal, a structure that lends itself readily to rapid cell growth. Measurements of the membrane’sviscosity show it as a fluid one hundred times as viscous as water, similar to a thin oil. The phospholipid molecules diffuse readily in the plane of the bilayer. Many of the membrane’s proteins also have this freedom of movement, but some are fixed in the membrane by interaction with the cell’s cytoskeleton. Newly synthesized phospholipids insert themselves easily into the existing cell membrane. Intrinsic proteins are inserted during their synthesis on ribosomes bound to the endoplasmic reticulum, whereas extrinsic proteins found on the internal surface of the cell membrane are synthesized on free, or unattached, ribosomes, liberated into the cytoplasm, and then brought to the membrane.
Ligand-gated ion channel: nicotinic acetylcholine receptor
The nicotinic acetylcholine receptor is an example of a ligand-gated ion channel. It is composed of five subunits arranged symmetrically around a central conducting pore. Upon binding acetylcholine, the channel opens and allows diffusion of sodium (Na+) and potassium (K+) ions through the conducting pore.
The chemical structure of the cell membrane makes it remarkably flexible, the ideal boundary for rapidly growing and dividing cells. Yet the membrane is also a formidable barrier, allowing some dissolved substances, or solutes, to pass while blocking others. Lipid-soluble molecules and some small molecules can permeate the membrane, but the lipid bilayer effectively repels the many large, water-soluble molecules and electrically charged ions that the cell must import or export in order to live. Transport of these vital substances is carried out by certain classes of intrinsic proteins that form a variety of transport systems: some are open channels, which allow ions to diffuse directly into the cell; others are “facilitators,” which, through a little-understood chemical transformation, help solutes diffuse past the lipid screen; yet others are “pumps,” which force solutes through the membrane when they are not concentrated enough to diffuse spontaneously. Particles too large to be diffused or pumped are often swallowed or disgorged whole by an opening and closing of the membrane.
Behind this movement of solutes across the cell membrane is the principle of diffusion. According to this principle, a dissolved substance diffuses down a concentration gradient; that is, given no energy from an outside source, it moves from a place where its concentration is high to a place where its concentration is low. Diffusion continues down this gradually decreasing gradient until a state of equilibrium is reached, at which point there is an equal concentration in both places and an equal, random diffusion in both directions.
A solute at high concentration is at high free energy; that is, it is capable of doing more “work” (the work being that of diffusion) than a solute at low concentration. In performing the work of diffusion, the solute loses free energy, so that, when it reaches equilibrium at a lower concentration, it is unable to return spontaneously (under its own energy) to its former high concentration. However, by the addition of energy from an outside source (through the work of an ion pump, for example), the solute may be returned to its former concentration and state of high free energy. This “coupling” of work processes is, in effect, a transferal of free energy from the pump to the solute, which is then able to repeat the work of diffusion. (See aboveCoupled chemical reactions.)
For most substances of biological interest, the concentrations inside and outside the cell are different, creating concentration gradients down which the solutes spontaneously diffuse, provided they can permeate the lipid bilayer. Membrane channels and diffusion facilitators bring them through the membrane by passive transport; that is, the changes that the proteins undergo in order to facilitate diffusion are powered by the diffusing solutes themselves. For the healthy functioning of the cell, certain solutes must remain at different concentrations on each side of the membrane; if through diffusion they approach equilibrium, they must be pumped back up their gradients by the process of active transport. Those membrane proteins serving as pumps accomplish this by coupling the energy required for transport to the energy produced by cell metabolism or by the diffusion of other solutes.
Permeation is the diffusion, through a barrier, of a substance in solution. The rates at which biologically important molecules cross the cell membrane through permeation vary over an enormous range. Proteins and sugar polymers do not permeate at all; in contrast, water and alcohols permeate most membranes in less than a second. This variation, caused by the lipid bilayer, gives the membrane its characteristic permeability. Permeability is measured as the rate at which a particular substance in solution crosses the membrane.
For all cell membranes that have been studied in the laboratory, permeability increases in parallel with the permeant’s ability to dissolve in organic solvents. The consistency of this parallel has led researchers to conclude that permeability is a function of the fatty acid interior of the lipid bilayer, rather than its phosphoryl exterior. This property of dissolving in organic solvents rather than water is given a unit of measure called the partition coefficient. The greater the solubility of a substance, the higher its partition coefficient, and the higher the partition coefficient, the higher the permeability of the membrane to that particular substance. For example, the water solubility of hydroxyl, carboxyl, and amino groups reduces their solubility in organic solvents and, hence, their partition coefficients. Cell membranes have been observed to have low permeability toward these groups. In contrast, lipid-soluble methyl residues and hydrocarbon rings, which have high partition coefficients, penetrate cell membranes more easily—a property useful in designing chemotherapeutic and pharmacological drugs.
For two molecules of the same partition coefficient, the one of greater molecular weight, or size, will in general cross the membrane more slowly. In fact, even molecules with very low partition coefficients can penetrate the membrane if they are small enough. Water, for example, is insoluble in organic solvents, yet it permeates cell membranes because of the small size of its molecules. The size selectivity of the lipid bilayer is a result of its being not a simple fluid, the molecules of which move around and past a diffusing molecule, but an organized matrix, a kind of fixed grate, composed of the fatty acid chains of the phospholipids through which the diffusing molecule must fit.
Many substances do not actually cross the cell membrane through permeation of the lipid bilayer. Some electrically charged ions, for example, are repelled by organic solvents and therefore cross cell membranes with great difficulty, if at all. In these cases special holes in the membrane, called channels, allow specific ions and small molecules to diffuse directly through the bilayer.
Diffusion of ions across a semipermeable membrane. (A) A high concentration of KCl is placed on side 1, opposite a semipermeable membrane from a low concentration. The membrane allows only K+ to diffuse, thereby establishing an electrical potential difference across the membrane. (B) The separation of charge creates an electrostatic voltage force, which draws some K+ back to side 1. (C) At equilibrium, there is no net flux of K+ in either direction. Side 1, with the higher concentration of KCl, has a negative charge compared with side 2.
Biophysicists measuring the electric current passing through cell membranes have found that, in general, cell membranes have a vastly greater electrical conductance than does a membrane bilayer composed only of phospholipids and sterols. This greater conductance is thought to be conferred by the cell membrane’s proteins. A current flowing across a membrane often appears on a recording instrument as a series of bursts of various heights. These bursts represent current flowing through open channels, which are merely holes formed by intrinsic proteins traversing the lipid bilayer. No significant current flows through the membrane when no channel is open; multiple bursts are recorded when more than one channel is open.
A rich variety of channels has been isolated and analyzed from a wide range of cell membranes. Invariably intrinsic proteins, they contain numerous amino acid sequences that traverse the membrane, clearly forming a specific hole, or pore. Certain channels open and close spontaneously. Some are gated, or opened, by the chemical action of a signaling substance such as calcium,acetylcholine, or glycine, whereas others are gated by changes in the electrical potential across the membrane. Channels may possess a narrow specificity, allowing passage of only potassium or sodium, or a broad specificity, allowing passage of all positively charged ions (cations) or of all negatively charged ions (anions). There are channels called gap junctions that allow the passage of molecules between pairs of cells (see belowThe cell matrix and cell-to-cell communication).
The gating of channels with a capacity for ion transport is the basis of the many nerve-nerve, nerve-muscle, and nerve-gland interactions underlying neurobiological behaviour. These actions depend on the electric potential of the cell membrane, which varies with the prevailing constituents in the cell’s environment. For example, if a channel that admits only potassium ions is present in a membrane separating two different potassium chloride solutions, the positively charged potassium ions tend to flow down their concentration gradient through the channel. The negatively charged chloride ions remain behind. This separation of electric charges sets up an electric potential across the membrane called the diffusion potential. The size of this potential depends on, among other factors, the difference in concentrations of the permeating ion across the membrane. The cell membrane in general contains the channels of widely different ion specificities, each channel contributing to the overall membrane potential according to the permeability and concentration ratio of the ion passing through it. Since the channels are often gated, the membrane’s potential is determined by which channels are open; this in turn depends on the concentrations of signaling molecules and may change with time according to the membrane potential itself.
Most cells have about a tenfold higher concentration of sodium ions outside than inside and a reverse concentration ratio of potassium ions. Free calcium ions can be 10,000 times more concentrated outside the cell than inside. Thus, sodium-, potassium-, and calcium-selective membrane channels, by allowing the diffusion of those ions past the cell membrane and causing fluctuations in the membrane’s electric potential, frequently serve as transmitters of signals from nerve cells. Ion diffusion threatens to alter the concentration of ions necessary for the cell to function. The proper distribution of ions is restored by the action of ion pumps (see belowPrimary active transport).
Many water-soluble molecules that cannot penetrate the lipid bilayer are too large to fit through open channels. In this category are sugars and amino acids. Some ions too do not diffuse through channels. These vital substances enter and leave the cell through the action of membrane transporters, which, like channels, are intrinsic proteins that traverse the cell membrane. Unlike channels, transporter molecules do not simply open holes in the membrane. Rather, they present sites on one side of the membrane to which molecules bind through chemical attraction. The binding site is highly specific, often fitting the atomic structure of only one type of molecule. When the molecule has attached to the binding site, then, in a process not fully understood, the transporter brings it through the membrane and releases it on the other side.
This action is considered a type of diffusion because the transported molecules move down their concentration gradients, from high concentration to low. To activate the action of the transporter, no other energy is needed than that of the chemical binding of the transported molecules. This action upon the transporter is similar to catalysis, except that the molecules (in this context called substrates) catalyze not a chemical reaction but their own translocation across the cell membrane. Two such substrates are glucose and the bicarbonate ion.
The glucose transporter
This sugar-specific transport system enables half of the glucose present inside the cell to leave within four seconds at normal body temperature. The glucose transporter is clearly not a simple membrane channel. First, unlike a channel, it does not select its permeants by size, as one type of glucose is observed to move through the system a thousand times faster than its identically sized optical isomer. Second, it operates much more slowly than do most channels, moving only 1,000 molecules per second while a channel moves 1,000,000 ions. The most important difference between a membrane channel and the glucose transporter is the conformational change that the transporter undergoes while moving glucose across the membrane. Alternating between two conformations, it moves its glucose-binding site from one side of the membrane to the other. By “flipping” between its two conformational states, the transporter facilitates the diffusion of glucose; that is, it enables glucose to avoid the barrier of the cell membrane while moving spontaneously down its concentration gradient. When the concentration reaches equilibrium, net movement of glucose ceases.
A facilitated diffusion system for glucose is present in many cell types. Similar systems transporting a wide range of other substrates (e.g., different sugars, amino acids, nucleosides, and ions) are also present.
The best-studied of the facilitated diffusion systems is that which catalyzes the exchange of anions across the red blood cell membrane. The exchange of hydroxyl for bicarbonate ions, each ion simultaneously being moved down its concentration gradient in opposite directions by the same transport molecule, is of great importance in enhancing the blood’s capacity to carry carbon dioxide from tissues to the lungs. The exchange molecule for these anions is the major intrinsic protein of red blood cells; one million of them are present on each cell, the polypeptide chain of each molecule traversing the membrane at least six times.
In some cases the problem of forcing a substrate up its concentration gradient is solved by coupling that upward movement to the downward flow of another substrate. In this way the energy-expending diffusion of the driving substrate powers the energy-absorbing movement of the driven substrate from low concentration to high. Because this type of active transport is not powered directly by the energy released in cell metabolism (see belowPrimary active transport), it is called secondary.
There are two kinds of secondary active transport: counter-transport, in which the two substrates cross the membrane in opposite directions, and cotransport, in which they cross in the same direction.
Counter-transport
An example of this system (also called antiport) begins with the sugar transporter described above. There are equal concentrations of glucose on both sides of the cell. A high concentration of galactose is then added outside the cell. Galactose competes with glucose for binding sites on the transport protein, so that mostly galactose—and a little glucose—enter the cell. The transporter itself, undergoing a conformational change, presents its binding sites for sugar at the inner face of the membrane. Here, at least transiently, glucose is in excess of galactose; it binds to the transporter and leaves the cell as the transporter switches back to its original conformation. Thus, glucose is pumped out of the cell against its gradient in exchange for the galactose riding into the cell down its own gradient.
Many counter-transport systems operate across the cell membranes of the body. A well-studied system (present in red blood cells, nerve cells, and muscle cells) pumps one calciumion out of the cell in exchange for two or three sodium ions. This system helps maintain the low calcium concentration required for effective cellular activity. A different system, present in kidney cells, counter-transports hydrogen ions and sodium ions in a one-for-one ratio. This is important in stabilizing acidity by transporting hydrogen ions out of the body as needed.
Co-transport
In co-transport (sometimes called symport) two species of substrate, generally an ion and another molecule or ion, must bind simultaneously to the transporter before its conformational change can take place. As the driving substrate is transported down its concentration gradient, it drags with it the driven substrate, which is forced to move up its concentration gradient. The transporter must be able to undergo a conformational change when not bound to either substrate, so as to complete the cycle and return the binding sites to the side from which driving and driven substrates both move.
Sodium ions are usually the driving substrates in the co-transport systems of animal cells, which maintain high concentrations of these ions through primary active transport. The driven substrates include a variety of sugars, amino acids, and other ions. During the absorption of nutrients, for example, sugars and amino acids are removed from the intestine by co-transport with sodium ions. After passing across the glomerular filter in the kidney, these substrates are returned to the body by the same system. Plant and bacterial cells usually use hydrogen ions as the driving substrate; sugars and amino acids are the most common driven substrates. When the bacteriumEscherichia coli must metabolize lactose, it co-transports hydrogen ions with lactose (which can reach a concentration 1,000 times higher than that outside the cell).
Human red blood cells contain a high concentration of potassium and a low concentration of sodium, yet the plasma bathing the cells is high in sodium and low in potassium. When whole blood is stored cold under laboratory conditions, the cells lose potassium and gain sodium until the concentrations across the membrane for both ions are at equilibrium. When the cells are restored to body temperature and given appropriate nutrition, they extrude sodium and take up potassium, transporting both ions against their respective gradients until the previous high concentrations are reached. This ion pumping is linked directly to the hydrolysis of adenosine triphosphate (ATP), the cell’s repository of metabolic energy (see aboveCoupled chemical reactions). For every molecule of ATP split, three ions of sodium are pumped out of the cell and two of potassium are pumped in.
An enzyme called sodium-potassium-activated ATPase has been shown to be the sodium-potassium pump, the protein that transports the ions across the cell membrane while splitting ATP. Widely distributed in the animal kingdom and always associated with the cell membrane, this ATPase is found at high concentration in cells that pump large amounts of sodium (e.g., in mammalian kidneys, in salt-secreting glands of marine birds, and in the electric organs of eels). The enzyme, an intrinsic protein, exists in two major conformations whose interconversion is driven by the splitting of ATP or by changes in the transmembrane flows of sodium and potassium. When only sodium is present in the cell, the inorganic phosphate split from ATP during hydrolysis is transferred to the enzyme. Release of the chemically bound phosphate from the enzyme is catalyzed by potassium. Thus, the complete action of ATP splitting has been demonstrated to require both sodium (to catalyze the transfer of the phosphate to the enzyme) and potassium (to catalyze the release of the phosphate and free the enzyme for a further cycle of ATP splitting). Apparently, only after sodium has catalyzed the transferal of the phosphate to the enzyme can it be transported from the cell. Similarly, only after potassium has released the phosphate from the enzyme can it be transported into the cell. This overall reaction, completing the cycle of conformational changes in the enzyme, involves a strict coupling of the splitting of ATP with the pumping of sodium and potassium. It is this coupling that creates primary active transport.
The sodium-potassium pump extrudes one net positive charge during each cycle of ATP splitting. This flow of current induces an electric potential across the membrane that adds to the potentials brought about by the diffusion of ions through gated channels. The pump’s contribution to the overall potential is important in certain specialized nerve cells.
Hydrochloric acid is produced in the stomach by the active transport of hydrogen ions from the blood across the stomach lining, or gastric mucosa. Hydrogen concentration gradients of nearly one million can be achieved by a hydrogen-potassium-activated ATP-splitting intrinsic protein in the cells lining the stomach. Apart from its specific ion requirements, the properties of this enzyme are remarkably similar to those of the sodium-potassium-activated enzyme and the calcium-activated enzyme. Other hydrogen-pumping ATP-splitting primary active transporters occur in intracellular organelles, in bacteria, and in plant cells (see belowThe mitochondrion and the chloroplast). The steep gradient of hydrogen ions represents a store of energy that can be harnessed to the accumulation of nutrients or, in the case of bacterial flagella, to the powering of cell movement.
In bringing about transmembrane movements of large molecules, the cell membrane itself undergoes concerted movements during which part of the fluid medium outside of the cell is internalized (endocytosis) or part of the cell’s internal medium is externalized (exocytosis). These movements involve a fusion between membrane surfaces, followed by the re-formation of intact membranes.
Endocytosis and exocytosis.
Endocytosis and exocytosis are fundamental to the process of intracellular digestion. Food particles are taken into the cell via endocytosis into a vacuole. Lysosomes attach to the vacuole and release digestive enzymes to extract nutrients. The leftover waste products of digestion are carried to the plasma membrane by the vacuole and eliminated through the process of exocytosis.
In this process the cell membrane engulfs portions of the external medium, forms an almost complete sphere around it, and then draws the membrane-bounded vesicle, called an endosome, into the cell. Several types of endocytosis have been distinguished: in pinocytosis, the vesicles are small and contain fluid; in phagocytosis, the vesicles are larger and contain solid matter; and in receptor-mediated endocytosis, material binds to a specific receptor on the external face of the cell membrane, triggering the process by which it is engulfed. Cholesterol enters cells by the last route.
Phagocytosis.
The process by which cells engulf solid matter is called phagocytosis. There are four essential steps in phagocytosis: (1) the plasma membrane entraps the food particle, (2) a vacuole forms within the cell to contain the food particle, (3) lysosomes fuse with the food vacuole, and (4) enzymes of the lysosomes digest the food particle.
Receptor-mediated endocytosis.
Receptors play key roles in many cellular processes. For example, receptor-mediated endocytosis enables cells to ingest molecules such as proteins that are necessary for normal cell functioning.
In exocytosis, material synthesized within the cell that has been packaged into membrane-bound vesicles is exported from the cell following the fusion of the vesicles with the external cell membrane. The materials so exported are cell-specific protein products, neurotransmitters, and a variety of other molecules.
The presence of internal membranes distinguishes eukaryotic cells (cells with a nucleus) from prokaryotic cells (those without a nucleus). Prokaryotic cells are small (one to five micrometres in length) and contain only a single cell membrane; metabolic functions are often confined to different patches of the membrane rather than to areas in the body of the cell. Typical eukaryotic cells, by contrast, are much larger, the cell membrane constituting only 10 percent or less of the total cellular membrane. Metabolic functions in these cells are carried out in the organelles, compartments sequestered from the cell body, or cytoplasm, by internal membranes.
This section discusses internal membranes as structural and functional components in the organelles and vesicles of eukaryotic cells. The principal organelles—the nucleus,mitochondrion, and (in plants)chloroplast—are discussed elsewhere (see belowThe nucleus;The mitochondrion and the chloroplast). Of the remaining organelles, the lysosomes, peroxisomes, and (in plants) glyoxysomes enclose extremely reactive by-products and enzymes. Internal membranes form the mazelike endoplasmic reticulum, where cell membrane proteins and lipids are synthesized, and they also form the stacks of flattened sacs called the Golgi apparatus, which is associated with the transport and modification of lipids, proteins, and carbohydrates. Finally, internal cell membranes can form storage and transport vesicles and the vacuoles of plant cells. Each membrane structure has its own distinct composition of proteins and lipids enabling it to carry out unique functions.
Like the cell membrane, membranes of some organelles contain transport proteins, or permeases, that allow chemical communication between organelles. Permeases in the lysosomal membrane, for example, allow amino acids generated inside the lysosome to cross into the cytoplasm, where they can be used for the synthesis of new proteins. Communication between organelles is also achieved by the membrane budding processes of endocytosis and exocytosis, which are essentially the same as in the cell membrane (see aboveTransport across the membrane). On the other hand, the biosynthetic and degradative processes taking place in different organelles may require conditions greatly different from those of other organelles or of the cytosol (the fluid part of the cell surrounding the organelles). Internal membranes maintain these different conditions by isolating them from one another. For example, the internal space of lysosomes is much more acidic than that of the cytosol—pH 5 as opposed to pH 7—and is maintained by specific proton-pumping transport proteins in the lysosome membrane.
Another function of organelles is to prevent competing enzymatic reactions from interfering with one another. For instance, essential proteins are synthesized on the rough endoplasmic reticulum and in the cytosol, while unwanted proteins are broken down in the lysosomes and also, to some extent, in the cytosol. Similarly, fatty acids are made in the cytosol and then either broken down in the mitochondria for the synthesis of ATP or degraded in the peroxisomes with concomitant generation of heat. These processes must be kept isolated. Organelle membranes also prevent potentially lethal by-products or enzymes from attacking sensitive molecules in other regions of the cell by sequestering such degradative activities in their respective membrane-bounded compartments.
Structural formula of cholesterol.
The internal membranes of eukaryotic cells differ both structurally and chemically from the outer cell membrane. Like the outer membrane, they are constructed of a phospholipid bilayer into which are embedded, or bound, specific membrane proteins (see aboveChemical composition and structure of the membrane). The three major lipids forming the outer membrane—phospholipids, cholesterol, and glycolipids—are also found in the internal membranes, but in different concentrations. Phospholipid is the primary lipid forming all cellular membranes. Cholesterol, which contributes to the fluidity and stability of all membranes, is found in internal membranes at about 25 percent of the concentration in the outer membrane. Glycolipids are found only as trace components of internal membranes, whereas they constitute approximately 5 percent of the outer membrane lipid.
Cellular organelles and their membranes
Cellular organelles and their membranes (B)
No text.
Plant cells contain membrane-bound organelles, including fluid-filled spaces, called vacuoles, that play an important role in maintaining the rigidity of a plant.
Most plant cells contain one or more membrane-bound vesicles called vacuoles. Within the vacuole is the cell sap, a water solution of salts and sugars kept at high concentration by the active transport of ions through permeases in the vacuole membrane. Proton pumps also maintain high concentrations of protons in the vacuole interior. These high concentrations cause the entry, via osmosis, of water into the vacuole, which in turn expands the vacuole and generates a hydrostatic pressure, called turgor, that presses the cell membrane against the cell wall. Turgor is the cause of rigidity in living plant tissue.
In the mature plant cell, as much as 90 percent of cell volume may be taken up by a single vacuole; immature cells typically contain several smaller vacuoles.
Potentially dangerous hydrolytic enzymes functioning in acidic conditions (pH 5) are segregated in the lysosomes to protect the other components of the cell from random destruction. Lysosomes are bound by a single phospholipid bilayer membrane. They vary in size and are formed by the fusion of Golgi-derived vesicles with endosomes derived from the cell surface. Enzymes known to be present in the lysosomes include hydrolases that degrade proteins, nucleic acids, lipids, glycolipids, and glycoproteins. Hydrolases are most active in the acidity maintained in the lysosomes. After the material is broken down, lipids and amino acids are transported across the lysosomal membrane by permeases for use in biosynthesis. The remaining debris generally stays within the lysosome and is called a residual body.
Microbodies are roughly spherical in shape, bound by a single membrane, and are usually 0.5 to 1 micrometre in diameter. There are several types, by far the most common of which is the peroxisome. Peroxisomes derive their name from hydrogen peroxide, a reactive intermediate in the process of molecular breakdown that occurs in the microbody. Peroxisomes contain type II oxidases, which are enzymes that use molecular oxygen in reactions to oxidize organic molecules. A product of these reactions is hydrogen peroxide, which is further metabolized into water and oxygen by the enzymecatalase, a predominant constituent of peroxisomes. In addition, peroxisomes contain other enzyme systems that degrade various lipids.
The plant glyoxysome is a peroxisome that also contains the enzymes of the glyoxylate cycle, which is crucial to the conversion of fat into carbohydrate.
The endoplasmic reticulum (ER) plays a major role in the biosynthesis of proteins. Proteins that are synthesized by ribosomes on the ER are transported into the Golgi apparatus for processing. Some of these proteins will be secreted from the cell, others will be inserted into the plasma membrane, and still others will be inserted into lysosomes.
The endoplasmic reticulum (ER) is a system of membranous cisternae (flattened sacs) extending throughout the cytoplasm. Often it constitutes more than half of the total membrane in the cell. This structure was first noted in the late 19th century, when studies of stained cells indicated the presence of some type of extensive cytoplasmic structure, then termed the gastroplasm. The electron microscope made possible the study of the morphology of this organelle in the 1940s, when it was given its present name.
The endoplasmic reticulum can be classified in two functionally distinct forms, the smooth endoplasmic reticulum (SER) and the rough endoplasmic reticulum (RER). The morphological distinction between the two is the presence of protein-synthesizing particles, called ribosomes, attached to the outer surface of the RER.
The functions of the SER, a meshwork of fine tubular membrane vesicles, vary considerably from cell to cell. One important role is the synthesis of phospholipids and cholesterol, which are major components of the plasma and internal membranes. Phospholipids are formed from fatty acids,glycerol phosphate, and other small water-soluble molecules by enzymes bound to the ER membrane with their active sites facing the cytosol. Some phospholipids remain in the ER membrane, where, catalyzed by specific enzymes within the membranes, they can “flip” from the cytoplasmic side of the bilayer, where they were formed, to the exoplasmic, or inner, side. This process ensures the symmetrical growth of the ER membrane. Other phospholipids are transferred through the cytoplasm to other membranous structures, such as the cell membrane and the mitochondrion, by special phospholipid transfer proteins.
In liver cells, the SER is specialized for the detoxification of a wide variety of compounds produced by metabolic processes. Liver SER contains a number of enzymes called cytochrome P450, which catalyze the breakdown of carcinogens and other organic molecules. In cells of the adrenal glands and gonads, cholesterol is modified in the SER at one stage of its conversion to steroid hormones. Finally, the SER in muscle cells, known as the sarcoplasmic reticulum, sequesters calcium ions from the cytoplasm. When the muscle is triggered by nerve stimuli, the calcium ions are released, causing muscle contraction.
The RER is generally a series of connected flattened sacs. It plays a central role in the synthesis and export of proteins and glycoproteins and is best studied in the secretory cells specialized in these functions. The many secretory cells in the human body include liver cells secreting serum proteins such as albumin,endocrine cells secreting peptide hormones such as insulin,salivary gland and pancreatic acinar cells secreting digestive enzymes, mammary gland cells secreting milk proteins, and cartilage cells secreting collagen and proteoglycans.
Ribosomes are particles that synthesize proteins from amino acids. They are composed of four RNA molecules and between 40 and 80 proteins assembled into a large and a small subunit. Ribosomes are either free (i.e., not bound to membranes) in the cytoplasm of the cell or bound to the RER. Lysosomal enzymes, proteins destined for the ER, Golgi, and cell membranes, and proteins to be secreted from the cell are among those synthesized on membrane-bound ribosomes. Fabricated on free ribosomes are proteins remaining in the cytosol and those bound to the internal surface of the outer membrane, as well as those to be incorporated into the nucleus, mitochondria, chloroplasts, peroxisomes, and other organelles. Special features of proteins label them for transport to specific destinations inside or outside of the cell. In 1971 German-born cellular and molecular biologist Günter Blobel and Argentinian-born cellular biologist David Sabatini suggested that the amino-terminal portion of the protein (the first part of the molecule to be made) could act as a “signal sequence.” They proposed that such a signal sequence would facilitate the attachment of the growing protein to the ER membrane and lead the protein either into the membrane or through the membrane into the ER lumen (interior).
The signal hypothesis has been substantiated by a large body of experimental evidence. Translation of the blueprint for a specific protein encoded in a messenger RNA molecule begins on a free ribosome. As the growing protein, with the signal sequence at its amino-terminal end, emerges from the ribosome, the sequence binds to a complex of six proteins and one RNA molecule known as the signal recognition particle (SRP). The SRP also binds to the ribosome to halt further formation of the protein. The membrane of the ER contains receptor sites that bind the SRP-ribosome complex to the RER membrane. Upon binding, translation resumes, with the SRP dissociating from the complex and the signal sequence and remainder of the nascent protein threading through the membrane, via a channel called a translocon, into the ER lumen. At that point, the protein is permanently segregated from the cytosol. In most cases, the signal sequence is cleaved from the protein by an enzyme called signal peptidase as it emerges on the luminal surface of the ER membrane. In addition, in a process known as glycosylation,oligosaccharide (complex sugar) chains are often added to the protein to form a glycoprotein. Inside the ER lumen, the protein folds into its characteristic three-dimensional conformation.
Within the lumen, proteins that will be secreted from the cell diffuse into the transitional portion of the ER, a region that is largely free of ribosomes. There the molecules are packaged into small membrane-bounded transport vesicles, which separate from the ER membrane and move through the cytoplasm to a target membrane, usually the Golgi complex. There the transport vesicle membrane fuses with the Golgi membrane, and the contents of the vesicle are delivered into the lumen of the Golgi. This, like all processes of vesicle budding and fusion, preserves the sidedness of the membranes; that is, the cytoplasmic surface of the membrane always faces outward, and the luminal contents are always sequestered from the cytoplasm.
Certain nonsecretory proteins made on the RER remain part of the membrane system of the cell. These membrane proteins have, in addition to the signal sequence, one or more anchor regions composed of lipid-soluble amino acids. The amino acids prevent passage of the protein completely into the ER lumen by anchoring it into the phospholipid bilayer of the ER membrane.
The Golgi complex is the site of the modification, completion, and export of secretory proteins and glycoproteins. This organelle, first described by the Italian cytologist Camillo Golgi in 1898, has a characteristic structure composed of five to eight flattened, disk-shaped, membrane-defined cisternae arranged in a stack. Secretory proteins and glycoproteins, cell membrane proteins and glycoproteins, lysosomal proteins, and some glycolipids all pass through the Golgi structure at some point in their maturation. In plant cells, much of the cell wall material passes through the Golgi as well.
The Golgi apparatus itself is structurally polarized, consisting of a “cis” face near the transitional region of the RER, a medial segment, and a “trans” face near the cell membrane. These faces are biochemically distinct, and the enzymatic content of each segment is markedly different. The cis face membranes are generally thinner than the others.
As the secretory proteins move through the Golgi, a number of chemical modifications may transpire. Important among these is the modification of carbohydrate groups. As described above, many secretory proteins are glycosylated in the ER. In the Golgi, specific enzymes modify the oligosaccharide chains of the glycoproteins by removing certain mannose residues and adding other sugars, such as galactose and sialic acid. These enzymes are known collectively as glycosidases and glycosyltransferases. Some secretory proteins will cease to be transported if their carbohydrate groups are modified incorrectly or not permitted to form. In some cases the carbohydrate groups are necessary for the stability or activity of the protein or for targeting the molecule for a specific destination.
Also within the Golgi or secretory vesicles are proteases that cut many secretory proteins at specific amino acid positions. This often results in activation of the secretory protein, an example being the conversion of inactive proinsulin to active insulin by removing a series of amino acids.
The release of proteins or other molecules from a secretory vesicle is most often stimulated by a nervous or hormonal signal. For example, a nerve cell impulse triggers the fusion of secretory vesicles to the membrane at the nerve terminal, where the vesicles release neurotransmitters into the synaptic cleft (the gap between nerve endings). The action is one of exocytosis: the vesicle and the cell membrane fuse, allowing the proteins and glycoproteins in the vesicle to be released to the cell exterior.
Synapse; neuron.
Chemical transmission of a nerve impulse at the synapse. The arrival of the nerve impulse at the presynaptic terminal stimulates the release of neurotransmitter into the synaptic gap. The binding of the neurotransmitter to receptors on the postsynaptic membrane stimulates the regeneration of the action potential in the postsynaptic neuron.
As secretory vesicles fuse with the cell membrane, the area of the cell membrane increases. Normal size is regained by the reuptake of membrane components through endocytosis. Regions bud in from the cell membrane and then fuse with internal membranes to effect recycling.
Not all proteins synthesized on the ER are destined for export. Many, such as the hydrolases in lysosomes, remain inside the cell; others become anchored in the membrane of internal organelles or in the cell membrane. It is presumed that each protein has some type of marker that fits a specific location in the cell.
Proteins synthesized on free ribosomes have segments that bind to specific receptors on the outer membrane of mitochondria, chloroplasts, or peroxisomes, allowing these proteins to be taken up only by these organelles. In the case of proteins synthesized in the RER, both the hydrolases destined for lysosomes and the secretory proteins are found initially in the same portion of the ER lumen. Studies have shown that these can be distinguished on the basis of their carbohydrate residues. The carbohydrate residues of lysosomal enzymes become modified in the cis-Golgi by the addition of certain phosphate groups. This critical modification allows the enzymes to bind to specific receptors on the membrane of the Golgi, which then directs them into vesicles leading to a lysosome rather than a secretory vesicle. In the lysosomes, proton pumps create an acidic environment that causes the release of the lysosomal enzyme from the membrane-bound receptors. Much of this sorting activity is mediated by coated vesicles containing the same fibrous outer protein, clathrin, used in endocytosis. These sorting vesicles also contain associated smaller proteins.
The nucleus is the information centre of the cell and is surrounded by a nuclear membrane in all eukaryotic organisms. It is separated from the cytoplasm by the nuclear envelope, and it houses the double-stranded, spiral-shaped deoxyribonucleic acid (DNA) molecules, which contain the genetic information necessary for the cell to retain its unique character as it grows and divides.
The presence of a nucleus distinguishes the eukaryotic cells of multicellular organisms from the prokaryotic, one-celled organisms such as bacteria. In contrast to the higher organisms, prokaryotes do not have nuclei, so their DNA is maintained in the same compartment as their other cellular components.
The primary function of the nucleus is the expression of selected subsets of the genetic information encoded in the DNA double helix. Each subset of a DNA chain, called a gene, codes for the construction of a specific protein out of a chain of amino acids. Information in DNA is not decoded directly into proteins, however. First it is transcribed, or copied, into a range of messenger ribonucleic acid (mRNA) molecules, each of which encodes the information for one protein (or more than one protein in bacteria). The mRNA molecules are then transported through the nuclear envelope into the cytoplasm, where they are translated, serving as templates for the synthesis of specific proteins.
The nucleus must not only synthesize the mRNA for many thousands of proteins, but it must also regulate the amounts synthesized and supplied to the cytoplasm. Furthermore, the amounts of each type of mRNA supplied to the cytoplasm must be regulated differently in each type of cell. In addition to mRNA, the nucleus synthesizes and exports other classes of RNA involved in the mechanisms of protein synthesis.
Structural organization of the nucleus
Structural organization of the nucleus (B)
No text.
During the first stages of cell division, the recognizable double-stranded chromosome is formed by two tightly coiled DNA strands (chromatids) joined at a point called the centromere. During the middle stage of cell division, the centromere duplicates, and the chromatid pair separates. Following cell division, the separated chromatids uncoil; the loosely coiled DNA, wrapped around its associated proteins (histones) to form beaded structures called nucleosomes, is termed chromatin.
Histone; nucleosome
DNA wrapped around clusters of histone proteins to form nucleosomes, which can coil to form solenoids.
The nucleus of the average human cell is only 6 micrometres (6 × 10−6 metre) in diameter, yet it contains about 1.8 metres of DNA. This is distributed among 46 chromosomes, each consisting of a single DNA molecule about 40 mm (1.5 inches) long. The extraordinary packaging problem this poses can be envisaged by a scale model enlarged a million times. On this scale a DNA molecule would be a thin string 2 mm thick, and the average chromosome would contain 40 km (25 miles) of DNA. With a diameter of only 6 metres, the nucleus would contain 1,800 km (1,118 miles) of DNA.
These contents must be organized in such a way that they can be copied into RNA accurately and selectively. DNA is not simply crammed or wound into the nucleus like a ball of string; rather, it is organized, by molecular interaction with specific nuclear proteins, into a precisely packaged structure. This combination of DNA with proteins creates a dense, compact fibre called chromatin. An extreme example of the ordered folding and compaction that chromatin can undergo is seen during cell division, when the chromatin of each chromosome condenses and is divided between two daughter cells (see belowCell division and growth).
Nucleosomes: the subunits of chromatin
The compaction of DNA is achieved by winding it around a series of small proteins called histones. Histones are composed of positively charged amino acids that bind tightly to and neutralize the negative charges of DNA. There are five classes of histone. Four of them, called H2A, H2B, H3, and H4, contribute two molecules each to form an octamer, an eight-part core around which two turns of DNA are wrapped. The resulting beadlike structure is called the nucleosome. The DNA enters and leaves a series of nucleosomes, linking them like beads along a string in lengths that vary between species of organism or even between different types of cell within a species. A string of nucleosomes is then coiled into a solenoid configuration by the fifth histone, called H1. One molecule of H1 binds to the site at which DNA enters and leaves each nucleosome, and a chain of H1 molecules coils the string of nucleosomes into the solenoid structure of the chromatin fibre.
Nucleosomes not only neutralize the charges of DNA, but they have other consequences. First, they are an efficient means of packaging. DNA becomes compacted by a factor of six when wound into nucleosomes and by a factor of about 40 when the nucleosomes are coiled into a solenoid chromatin fibre. The winding into nucleosomes also allows some inactive DNA to be folded away in inaccessible conformations, a process that contributes to the selectivity of gene expression.
Organization of chromatin fibre
Several studies indicate that chromatin is organized into a series of large radial loops anchored to specific scaffold proteins. Each loop consists of a chain of nucleosomes and may be related to units of genetic organization. This radial arrangement of chromatin loops compacts DNA about a thousandfold. Further compaction is achieved by a coiling of the entire looped chromatin fibre into a dense structure called a chromatid, two of which form the chromosome. During cell division, this coiling produces a 10,000-fold compaction of DNA.
The nuclear envelope is a double membrane composed of an outer and an inner phospholipid bilayer. The thin space between the two layers connects with the lumen of the rough endoplasmic reticulum (RER), and the outer layer is an extension of the outer face of the RER.
The inner surface of the nuclear envelope has a protein lining called the nuclear lamina, which binds to chromatin and other contents of the nucleus. The entire envelope is perforated by numerous nuclear pores. These transport routes are fully permeable to small molecules up to the size of the smallest proteins, but they form a selective barrier against movement of larger molecules. Each pore is surrounded by an elaborate protein structure called the nuclear pore complex, which selects molecules for entrance into the nucleus. Entering the nucleus through the pores are the nucleotide building blocks of DNA and RNA, as well as adenosine triphosphate, which provides the energy for synthesizing genetic material. Histones and other large proteins must also pass through the pores. These molecules have special amino acid sequences on their surface that signal admittance by the nuclear pore complexes. The complexes also regulate the export from the nucleus of RNA and subunits of ribosomes.
DNA in prokaryotes is also organized in loops and is bound to small proteins resembling histones, but these structures are not enclosed by a nuclear membrane.
Several features are common to the genetic structure of most organisms. First is the double-stranded DNA. Each strand of this molecule is a series of nucleotides, and each nucleotide is composed of a sugar-phosphate compound attached to one of four nitrogen-containing bases. The sugar-phosphate compounds link together to form the backbone of the strand. Each of the bases strung along the backbone is chemically attracted to a corresponding base on the parallel strand of the DNA molecule. This base pairing joins the two strands of the molecule much as rungs join the two sides of a ladder, and the chemical bonding of the base pairs twists the doubled strands into a spiral, or helical, shape.
The four nucleotide bases are adenine,cytosine,guanine, and thymine. DNA is composed of millions of these bases strung in an apparently limitless variety of sequences. It is in the sequence of bases that the genetic information is contained, each sequence determining the sequence of amino acids to be connected into proteins. A nucleotide sequence sufficient to encode one protein is called a gene. Genes are interspersed along the DNA molecule with other sequences that do not encode proteins. Some of these so-called untranslated regions regulate the activity of the adjacent genes, for example, by marking the points at which enzymes begin and cease transcribing DNA into RNA (see belowGenetic expression through RNA).
Rearrangements and modifications of the nucleotide sequences in DNA are exceptions to the rules of genetic expression and sometimes cause significant changes in the structure and function of cells. Different cells of the body owe their specialized structures and functions to different genes. This does not mean that the set of genetic information varies among the cells of the body. Indeed, for each cell the entire DNA content of the chromosomes is usually duplicated exactly from generation to generation, and, in general, the genetic content and arrangement is strikingly similar among different cell types of the same organism. As a result, the differentiation of cells can occur without the loss or irreversible inactivation of unnecessary genes, an observation that is reinforced by the presence of specific genes in a range of adult tissues. For example, normal copies of the genes encoding hemoglobin are present in the same numbers in red blood cells, which make hemoglobin, as in a range of other types of cells,
Antibody structure.
The structure of an antibody molecule represents the dramatic rearrangements of DNA that occur in the immune systems of mammals. Each antibody contains a light chain and a heavy chain that are encoded by different segments of DNA. These segments are subject to considerable variation and are thus able to produce many different antibodies.
.
Despite the general uniformity of genetic content in all the cells of an organism, studies have shown a few clear examples in some organisms of programmed, reversible change in the DNA of developing tissues. One of the most dramatic rearrangements of DNA occurs in the immune systems of mammals. The body’s defense against invasion by foreign organisms involves the synthesis of a vast range of antibodies by lymphocytes (a type of white blood cell). Antibodies are proteins that bind to specific invading molecules or organisms and either inactivate them or signal their destruction. The binding sites on each antibody molecule are formed by one light and one heavy amino acid chain, which are encoded by different segments of the DNA in the lymphocyte nucleus. These DNA segments undergo considerable rearrangements, resulting in the synthesis of a great variety of antibodies. Some invasive organisms, such as trypanosome parasites, which cause sleeping sickness, go to great lengths to rearrange their own DNA to evade the versatility of their hosts’ antibody production. The parasites are covered by a thick coat of glycoprotein (a protein with sugars attached). Given time, host organisms can overcome infection by producing antibodies to the parasites’ glycoprotein coat, but this reaction is anticipated and evaded by the selective rearrangement of the trypanosomes’ DNA encoding the glycoprotein, thus constantly changing the surface presented to the hosts’ immune system.
Careful comparisons of gene structure have also revealed epigenetic modifications, heritable changes that occur on the sugar-phosphate side of bases in the DNA and thus do not cause rearrangements in the DNA sequence itself. An example of an epigenetic modification involves the addition of a methyl group to cytosine bases. This appears to cause the inactivation of genes that do not need to be expressed in a particular type of cell. An important feature of the methylation of cytosine lies in its ability to be copied, so that methyl groups in a dividing cell’s DNA will result in methyl groups in the same positions in the DNA of both daughter cells.
The transcription of the genetic code from DNA to RNA, and the translation of that code from RNA into protein, exerts the greatest influence on the modulation of genetic information. The process of genetic expression takes place over several stages, and at each stage is the potential for further differentiation of cell types.
As explained above, genetic information is encoded in the sequences of the four nucleotide bases making up a DNA molecule. One of the two DNA strands is transcribed exactly into messenger RNA (mRNA), with the exception that the thymine base of DNA is replaced by uracil. RNA also contains a slightly different sugar component (ribose) from that of DNA (deoxyribose) in its connecting sugar-phosphate chain. Unlike DNA, which is stable throughout the cell’s life and of which individual strands are even passed on to many cell generations, RNA is unstable. It is continuously broken down and replaced, enabling the cell to change its patterns of protein synthesis.
Apart from mRNA, which encodes proteins, other classes of RNA are made by the nucleus. These include ribosomal RNA (rRNA), which forms part of the ribosomes and is exported to the cytoplasm to help translate the information in mRNA into proteins. Ribosomal RNA is synthesized in a specialized region of the nucleus called the nucleolus, which appears as a dense area within the nucleus and contains the genes that encode rRNA. This is also the site of assembly of ribosome subunits from rRNA and ribosomal proteins. Ribosomal proteins are synthesized in the cytoplasm and transported to the nucleus for subassembly in the nucleolus. The subunits are then returned to the cytoplasm for final assembly. Another class of RNA synthesized in the nucleus is transfer RNA (tRNA), which serves as an adaptor, matching individual amino acids to the nucleotide triplets of mRNA during protein synthesis.
The synthesis of RNA is performed by enzymes called RNA polymerases. In higher organisms there are three main RNA polymerases, designated I, II, and III (or sometimes A, B, and C). Each is a complex protein consisting of many subunits. RNA polymerase I synthesizes three of the four types of rRNA (called 18S, 28S, and 5.8S RNA); therefore it is active in the nucleolus, where the genes encoding these rRNA molecules reside. RNA polymerase II synthesizes mRNA, though its initial products are not mature RNA but larger precursors, called heterogeneous nuclear RNA, which are completed later (see belowProcessing of mRNA). The products of RNA polymerase III include tRNA and the fourth RNA component of the ribosome, called 5S RNA.
All three polymerases start RNA synthesis at specific sites on DNA and proceed along the molecule, linking selected nucleotides sequentially until they come to the end of the gene and terminate the growing chain of RNA. Energy for RNA synthesis comes from high-energy phosphate linkages contained in the nucleotide precursors of RNA. Each unit of the final RNA product is essentially a sugar, a base, and one phosphate, but the building material consists of a sugar, a base, and three phosphates. During synthesis two phosphates are cleaved and discarded for each nucleotide that is incorporated into RNA. The energy released from the phosphate bonds is used to link the nucleotides. The crucial feature of RNA synthesis is that the sequence of nucleotides joined into a growing RNA chain is specified by the sequence of nucleotides in the DNA template: each adenine in DNA specifies uracil in RNA, each cytosine specifies guanine, each guanine specifies cytosine, and each thymine in DNA specifies adenine. In this way the information encoded in each gene is transcribed into RNA for translation by the protein-synthesizing machinery of the cytoplasm.
In addition to specifying the sequence of amino acids to be polymerized into proteins, the nucleotide sequence of DNA contains supplementary information. For example, short sequences of nucleotides determine the initiation site for each RNA polymerase, specifying where and when RNA synthesis should occur. In the case of RNA polymerases I and II, the sequences specifying initiation sites lie just ahead of the genes. In contrast, the equivalent information for RNA polymerase III lies within the gene—that is, within the region of DNA to be copied into RNA. The initiation site on a segment of DNA is called a promoter. The promoters of different genes have some nucleotide sequences in common, but they differ in others. The differences in sequence are recognized by specific proteins called transcription factors, which are necessary for the expression of particular types of genes. The specificity of transcription factors contributes to differences in the gene expression of different types of cells.
During and after synthesis, mRNA precursors undergo a complex series of changes before the mature molecules are released from the nucleus. First, a modified nucleotide is added to the start of the RNA molecule by a reaction called capping. This cap later binds to a ribosome in the cytoplasm. The synthesis of mRNA is not terminated simply by the RNA polymerase’s detachment from DNA, but by chemical cleavage of the RNA chain. Many (but not all) types of mRNA have a simple polymer of adenosine residues added to their cleaved ends.
In addition to these modifications of the termini, startling discoveries in 1977 revealed that portions of newly synthesized RNA molecules are cut out and discarded. In many genes, the regions coding for proteins are interrupted by intervening sequences of nucleotides called introns. These introns must be excised from the RNA copy before it can be released from the nucleus as a functional mRNA. The number and size of introns within a gene vary greatly, from no introns at all to more than 50. The sum of the lengths of these intervening sequences is sometimes longer than the sum of the regions coding for proteins.
The removal of introns, called RNA splicing, appears to be mediated by small nuclear ribonucleoprotein particles (snRNP’s). These particles have RNA sequences that are complementary to the junctions between introns and adjacent coding regions. By binding to the junction ends, an snRNP twists the intron into a loop. It then excises the loop and splices the coding regions.
Although all the cell nuclei of an organism generally carry the same genes, there are conspicuous differences between the specialized cell types of the body. The source of these differences lies not so much in the occasional modification of DNA, as outlined above, but in the selective expression of DNA through RNA; in particular, it can be traced to processes regulating the amounts and activities of mRNA both during and after its synthesis in the nucleus.
The first level of regulation is mediated by variations in chromatin structure. In order to be transcribed, a gene must be assembled into a structurally distinct form of active chromatin. A second level of regulation is achieved by varying the frequency with which a gene in the active conformation is transcribed into RNA by an RNA polymerase. There is evidence for regulation of RNA synthesis at both these levels—for example, in response to hormoneinduction. At both levels, protein factors are believed to perform the regulation—for example, by binding to special promoter DNA regions flanking the transcribed gene.
After synthesis, RNA molecules undergo selective processing, which results in the export of only a subpopulation of RNA molecules to the cytoplasm. Furthermore, the stability in the cytoplasm of a particular type of mRNA can be regulated. For example, the hormoneprolactin increases synthesis of milk proteins in tissue by causing a twofold rise in the rate of mRNA synthesis; but it also causes a 17-fold rise in mRNA lifetime, so that in this case the main cause of increased protein synthesis is the prolonged availability of mRNA. Conversely, there is evidence for selective destabilization of some mRNA—such as histone mRNA, which is rapidly broken down when DNA replication is interrupted. Finally, there are many examples of selective regulation of the translation of mRNA into protein.
The internal membrane of a mitochondrion is elaborately folded into structures known as cristae. Cristae increase the surface area of the inner membrane, which houses the components of the electron-transport chain. Proteins known as F1F0ATPases that produce the majority of ATP used by cells are found throughout the cristae.
Chloroplast.
Internal structures of the chloroplast. The interior contains flattened sacs of photosynthetic membranes (thylakoids) formed by the invagination and fusion of the inner membrane. Thylakoids are usually arranged in stacks (grana) and contain the photosynthetic pigment (chlorophyll). The grana are connected to other stacks by simple membranes (lamellae) within the stroma, the fluid proteinaceous portion containing the enzymes essential for the photosynthetic dark reaction, or Calvin cycle.
Mitochondria and chloroplasts are the powerhouses of the cell. Mitochondria appear in both plant and animal cells as elongated cylindrical bodies, roughly one micrometre in length and closely packed in regions actively using metabolic energy. Mitochondria oxidize the products of cytoplasmic metabolism to generate adenosine triphosphate (ATP), the energy currency of the cell. Chloroplasts are the photosynthetic organelles in plants and some algae. They trap light energy and convert it partly into ATP but mainly into certain chemically reduced molecules that, together with ATP, are used in the first steps of carbohydrate production. Mitochondria and chloroplasts share a certain structural resemblance, and both have a somewhat independent existence within the cell, synthesizing some proteins from instructions supplied by their own DNA.
Both organelles are bounded by an external membrane that serves as a barrier by blocking the passage of cytoplasmic proteins into the organelle. An inner membrane provides an additional barrier that is impermeable even to small ions such as protons. The membranes of both organelles have a lipid bilayer construction (see aboveChemical composition and membrane structure). Located between the inner and outer membranes is the intermembrane space.
In mitochondria the inner membrane is elaborately folded into structures called cristae that dramatically increase the surface area of the membrane. In contrast, the inner membrane of chloroplasts is relatively smooth. However, within this membrane is yet another series of folded membranes that form a set of flattened, disklike sacs called thylakoids. The space enclosed by the inner membrane is called the matrix in mitochondria and the stroma in chloroplasts. Both spaces are filled with a fluid containing a rich mixture of metabolic products, enzymes, and ions. Enclosed by the thylakoid membrane of the chloroplast is the thylakoid space. The extraordinary chemical capabilities of the two organelles lie in the cristae and the thylakoids. Both membranes are studded with enzymatic proteins either traversing the bilayer or dissolved within the bilayer. These proteins contribute to the production of energy by transporting material across the membranes and by serving as electron carriers in important oxidation-reduction reactions.
Crucial to the function of mitochondria and chloroplasts is the chemistry of the oxidation-reduction, or redox, reaction. This controlled burning of material comprises the transfer of electrons from one compound, called the donor, to another, called the acceptor. All compounds taking part in redox reactions are ranked in a descending scale according to their ability to act as electron donors. Those higher in the scale donate electrons to their fellows lower down, which have a lesser tendency to donate, but a correspondingly greater tendency to accept, electrons. Each acceptor in turn donates electrons to the next compound down the scale, forming a donor-acceptor chain extending from the greatest donating ability to the least.
At the top of the scale is hydrogen, the most abundant element in the universe. The nucleus of a hydrogen atom is composed of one positively charged proton; around the nucleus revolves one negatively charged electron. In the atmosphere two hydrogen atoms join to form a hydrogen molecule (H2). In solution the two atoms pull apart, dissociating into their constituent protons and electrons. In the redox reaction the electrons are passed from one reactant to another. The donation of electrons is called oxidation, and the acceptance is called reduction—hence the descriptive term oxidation-reduction, indicating that one action never takes place without the other.
A hydrogen atom has a great tendency to transfer an electron to an acceptor. An oxygen atom, in contrast, has a great tendency to accept an electron. The burning of hydrogen by oxygen is, chemically, the transfer of an electron from each of two hydrogen atoms to oxygen, so that hydrogen is oxidized and oxygen reduced. The reaction is extremely exergonic; i.e., it liberates much free energy as heat. This is the reaction that takes place within mitochondria but is so controlled that the heat is liberated not at once but in a series of steps. The free energy, harnessed by the organelle, is coupled to the synthesis of ATP from adenosine diphosphate (ADP) and inorganic phosphate (Pi).
An analogy can be drawn between this controlled reaction and the flow of river water down a lock system. Without the locks, water flow would be rapid and uncontrolled, and no ship could safely ply the river. The locks force water to flow in small controlled steps conducive to safe navigation. But there is more to a lock system than this. The flow of water down the locks can also be harnessed to raise a ship from a lower to a higher level, with the water rather than the ship expending the energy. In mitochondria the burning of hydrogen is broken into a series of small indirect steps following the flow of electrons along a chain of donor-acceptors. Energy is funneled into the chemical bonding of ADP and Pi, raising the free energy of these two compounds to the high level of ATP.
The mitochondrion
The mitochondrion (B) Formation of the electron donors NADH and FADH2
NADH and FADH2 are compounds of high electron-donating capacity. Were they to transfer their electrons directly to oxygen, the resulting combustion would release a lethal burst of heat energy. Instead, the energy is released in a series of electron donor-acceptor reactions carried out within the cristae of the mitochondrion by a number of proteins and coenzymes that make up the electron-transport, or respiratory, chain.
The electron-transport chain
The proteins of this chain are embedded in the cristae membrane, actually traversing the lipid bilayer and protruding from the inner and outer surfaces. The coenzymes are dissolved in the lipid and diffuse through the membrane or across its surface. The proteins are arranged in three large complexes, each composed of a number of polypeptide chains. Each complex is, to continue the hydraulic analogy, a lock in the waterfall of the electron flow and the site at which energy from the overall redox reaction is tapped. The first complex, NADH dehydrogenase, accepts a pair of electrons from the primary electron donor NADH and is reduced in the process. It in turn donates these electrons to the coenzyme ubiquinone, a lipid-soluble molecule composed of a substituted benzene ring attached to a hydrocarbon tail. Ubiquinone, diffusing through the lipid of the cristae membrane, reaches the second large complex of the electron-transport chain, the b-c2 complex, which accepts the electrons, oxidizing ubiquinone and being itself reduced. (This complex can also accept electrons from the second primary electron donor, FADH2, a molecule below NADH in the electron-donating scale.) The b-c2 complex transfers the pair of electrons to cytochrome c, a small protein situated on the outer surface of the cristae membrane. From cytochrome c, electrons pass (four at a time) to the third large complex, cytochrome oxidase, which, in the final step of the chain, transfers the four electrons to two oxygen atoms and two protons, generating two water molecules.
Electron-transport chain.
The electron-transport chain embedded in the inner membrane of a mitochondrion is made up of a series of electron donors and electron acceptors. The transport of electrons begins with the acceptance of electrons by NADH dehydrogenase from NADH. The electrons are then passed to ubiquinone (coenzyme Q; site I), which carries them to the b-c2 complex. The electrons are then transferred to cytochrome c (site II), to cytochrome oxidase (site III), and finally to oxygen.
This transfer of electrons, from member to member of the electron-transport chain, provides energy for the synthesis of ATP through an indirect route. At the beginning of the electron-transport chain, NADH and FADH2 split hydrogen atoms into protons and electrons, transferring the electrons to the next protein complex and releasing the protons into the mitochondrial matrix. When each protein complex in turn transfers the electrons down the chain, it uses the energy released in this process to pump protons across the inner membrane into the intermembrane space. (For the dynamics of this pumping action, see aboveTransport across the membrane.) This transport of positively charged protons into the intermembrane space, opposite the negatively charged electrons in the matrix, creates an electrical potential that tends to draw the protons back across the membrane. A high concentration of protons outside the membrane also creates the conditions for their diffusion back into the matrix. However, as explained above, the inner membrane is extremely impermeable to protons. In order for the protons to flow back down the electrochemical gradient, they must traverse the membrane through transport molecules similar to the protein complexes of the electron-transfer chain. These molecules are the so-called F1F0ATPase, a complex protein that, transporting protons back into the matrix, uses the energy released to synthesize ATP. The protons then join the electrons and oxygen atoms to form water. (For further discussion of ATP production, see aboveCoupled chemical reactions.)
This complex chain of events, the basis of the cell’s ability to derive ATP from metabolic oxidation, was conceived in its entirety by the British biochemist Peter Mitchell in 1961. The years following the announcement of his chemiosmotic theory saw its ample substantiation and revealed its profound implications for cell biology.
The chemiosmotic theory
The four postulates of the chemiosmotic theory, including examples of their experimental substantiation, are as follows:
(1) The inner mitochondrial membrane is impermeable to protons,hydroxide ions, and other cations and anions. This postulate was validated when it was shown that substances allowing protons to flow readily across mitochondrial membranes uncouple oxidative electron transport from ATP production.
(2) Transfer of electrons down the electron-transport chain brings about pumping of protons across the inner membrane, from matrix to intermembrane space. This was demonstrated in laboratory experiments that reconstituted the components of the electron-transport chain in artificial membrane vesicles. The stimulation of electron transport caused a measurable buildup of protons within the vesicle.
(3) The flow of protons down a built-up electrochemical gradient occurs through a proton-dependent ATPase, so that ATP is synthesized from ADP and Pi whenever protons move through the enzyme. This hypothesis was confirmed by the discovery of what came to be known as the F1F0ATPase . Shaped like a knob attached to the membrane by a narrow stalk, F1F0ATPase covers the inner surface of the cristae. Its stalk (the F0 portion) penetrates the lipid bilayer of the inner membrane and is capable of catalyzing the transport of protons. The knob (the F1 portion) is capable of synthesizing as well as splitting, or hydrolyzing, ATP. F1F0ATPase is therefore reversible, either using the energy of proton diffusion to combine ADP and Pi or using the energy of ATP hydrolysis to pump protons out of the matrix.
(4) The inner membrane of the mitochondrion possesses a complement of proteins that brings about the transport of essential metabolites. Numerous carrier systems have been demonstrated to transport into the mitochondrion the products of metabolism that are transformed into substrates for the electron-transport chain. Best known is the ATP-ADP exchange carrier of the inner membrane. Neither ATP nor ADP, being large charged molecules, can cross the membrane unaided, but ADP must enter and ATP must leave the mitochondrial matrix in order for ATP synthesis to continue. A single protein conducts the counter-transport of ATP against ADP, the energy released by the flow of ATP down its concentration gradient being coupled to the pumping of ADP up its gradient and into the mitochondrion.
Light travels as packets of energy known as photons and is absorbed in this form by light-absorbing chlorophyll molecules embedded in the thylakoid membrane of the chloroplast. The chlorophyll molecules are grouped into antenna complexes, clusters of several hundred molecules that are anchored onto the thylakoid membrane by special proteins. Within each antenna complex is a specialized set of proteins and chlorophyll molecules that form a reaction centre. Photons absorbed by the other chlorophylls of the antenna are funneled into the reaction centre. The energy of the photon is absorbed by an electron of the reaction centre molecule in sufficient quantity to enable its acceptance by a nearby coenzyme, which cannot accept electrons at low energy levels. This coenzyme has a high electron-donor capability; it initiates the transfer of the electron down an electron-transport chain similar to that of the mitochondrion. Meanwhile, the loss of the negatively charged electron leaves a positively charged “hole” in the reaction centre chlorophyll molecule. This hole is filled by the enzymatic splitting of water into molecular oxygen, protons, and electrons and the transfer of an electron to the chlorophyll. The oxygen is released by the chloroplast, making its way out of the plant and into the atmosphere. The protons, in a process similar to that in the mitochondrion, are pumped through the thylakoid membrane and into the thylakoid space. Their facilitated diffusion back into the stroma through proteins embedded in the membrane powers the synthesis of ATP. This part of the photosynthetic process is called photosystem II.
At the end of the electron-transport chain in the thylakoid membrane is another reaction centre molecule. The electron is again energized by photons and then transported down another chain, which makes up photosystem I. This system uses the energy released in electron transfer to join a proton to nicotinamide adenine dinucleotide phosphate (NADP+), a phosphorylated derivative of NAD+, forming NADPH. NADPH is a high-energy electron donor that, with ATP, fuels the conversion of carbon dioxide into the carbohydrate foods of the plant cell.
Photosynthesis.
In photosynthesis, plants consume carbon dioxide and water and produce glucose and oxygen. Energy for this process is provided by light, which is absorbed by pigments, primarily chlorophyll. Chlorophyll is the pigment that gives plants their green colour.
NADPH remains within the stroma of the chloroplast for use in the fixation of carbon dioxide (CO2) during the Calvin cycle. In a complex cycle of chemical reactions, CO2 is bound to a five-carbon ribulose biphosphate compound. The resulting six-carbon intermediate is then split into three-carbon phosphoglycerate. With energy supplied by the breakdown of NADPH and ATP, this compound is eventually formed into glyceraldehyde 3-phosphate, an important sugar intermediate of metabolism. One glyceraldehyde molecule is exported from the chloroplast, for further conversion in the cytoplasm, for every five that undergo an ATP-powered re-formation into the five-carbon ribulose biphosphate. In this way three molecules of CO2 yield one molecule of glyceraldehyde 3-phosphate, while the entire fixation cycle hydrolyzes nine molecules of ATP and oxidizes six molecules of NADPH.
Calvin cycle.
Pathway of carbon dioxide fixation and reduction in photosynthesis, the Calvin cycle. The diagram represents one complete turn of the cycle, with the net production of one molecule of glyceraldehyde-3-phosphate (Gal3P). This three-carbon sugar phosphate usually is converted to either sucrose or starch.
The mitochondrion and chloroplast as independent entities
The mitochondrion and chloroplast as independent entities (B)
In addition to their remarkable metabolic capabilities, both mitochondria and chloroplasts synthesize on their own a number of proteins and lipids necessary for their structure and activity. Not only do they contain the machinery necessary for this, but they also possess the genetic material to direct it. DNA within these organelles has a circular structure reminiscent of prokaryotic, not eukaryotic, DNA. Also as in prokaryotes, the DNA is not associated with histones. Along with the DNA are protein-synthesizing ribosomes, of prokaryotic rather than eukaryotic size.
Only a small portion of the mitochondrion’s total number of proteins is synthesized within the organelle. Numerous proteins are encoded and made in the cytoplasm specifically for export into the mitochondrion. The mitochondrial DNA itself encodes only 13 different proteins. The proteins that contain subunits synthesized within the mitochondrion often also possess subunits synthesized in the cytoplasm. Mitochondrial and chloroplastic proteins synthesized in the cytoplasm have to enter the organelle by a complex process, crossing both the outer and the inner membranes. These proteins contain specific arrangements of amino acids known as leader sequences that are recognized by receptors on the outer membranes of the organelles. The proteins are then guided through membrane channels in an energy-requiring process.
Mitochondria and chloroplasts are self-dividing; they contain their own DNA and protein-synthesizing machinery, similar to that of prokaryotes. Chloroplasts produce ATP and trap photons by mechanisms that are complex and yet similar to those of certain prokaryotes. These phenomena have led to the theory that the two organelles are direct descendants of prokaryotes that entered primitive nucleated cells. Among billions of such events, a few could have led to the development of stable, symbiotic associations between nucleated hosts and prokaryotic parasites. The hosts would provide the parasites with a stable osmoticenvironment and easy access to nutrients, and the parasites would repay the hosts by providing an oxidative ATP-producing system or a photosynthetic energy-producing reaction.
The cytoskeleton is the name given to the fibrous network formed by different types of long protein filaments present throughout the cytoplasm of eukaryotic cells (cells containing a nucleus). The filaments of the cytoskeleton create a scaffold, or framework, that organizes other cell constituents and maintains the shape of the cell. In addition, some filaments cause coherent movements, both of the cell itself and of its internal organelles. Prokaryotic (nonnucleated) cells, which are generally much smaller than eukaryotic cells, contain a unique related set of filaments but, with few exceptions, do not possess true cytoskeletons. Their shapes and the shapes of certain eukaryotes, primarily yeast and other fungi, are determined by the rigid cell wall on the outside of the cell.
Four major types of cytoskeletal filaments are commonly recognized: actin filaments, microtubules, intermediate filaments, and septins. Actin filaments and microtubules are dynamic structures that continuously assemble and disassemble in most cells. Intermediate filaments are stabler and seem to be involved mainly in reinforcing cell structures, especially the position of the nucleus and the junctions that connect cells. Septins are involved in cell division and have been implicated in other cell functions. A wide variety of accessory proteins works in concert with each type of filament, linking filaments to one another and to the cell membrane and helping to form the networks that endow the cytoskeleton with its unique functions. Many of these accessory proteins have been characterized, revealing a rich diversity in the structure and function of the cytoskeleton.
Actin is a globular protein that polymerizes (joins together many small molecules) to form long filaments. Because each actin subunit faces in the same direction, the actin filament is polar, with different ends, termed “barbed” and “pointed.” An abundant protein in nearly all eukaryotic cells, actin has been extensively studied in muscle cells. In muscle cells, the actin filaments are organized into regular arrays that are complementary with a set of thicker filaments formed from a second protein called myosin. These two proteins create the force responsible for muscle contraction. When the signal to contract is sent along a nerve to the muscle, the actin and myosin are activated. Myosin works as a motor, hydrolyzing adenosine triphosphate (ATP) to release energy in such a way that a myosin filament moves along an actin filament, causing the two filaments to slide past each other. The thin actin filaments and the thick myosin filaments are organized in a structure called the sarcomere, which shortens as the filaments slide over one another. Skeletal muscles are composed of bundles of many long muscle cells; when the sarcomeres contract, each of these giant muscle cells shortens, and the overall effect is the contraction of the entire muscle. Although the stimulation pathways differ, heart muscle and smooth muscle (found in many internal organs and blood vessels) contract by a similar sliding filament mechanism.
Striated muscle; human biceps muscle.
The structure of striated muscle. Striated muscle tissue, such as the tissue of the human biceps muscle, consists of long, fine fibres, each of which is in effect a bundle of finer myofibrils. Within each myofibril are filaments of the proteins myosin and actin; these filaments slide past one another as the muscle contracts and expands. On each myofibril, regularly occurring dark bands, called Z lines, can be seen where actin and myosin filaments overlap. The region between two Z lines is called a sarcomere; sarcomeres can be considered the primary structural and functional unit of muscle tissue.
Actin is also present in non-muscle cells, where it forms a meshwork of filaments responsible for many types of cellular movement. The meshwork consists of actin filaments that are attached to the cell membrane and to each other. The length of the filaments and the architecture of their attachments determine the shape and consistency of a cell. A large number of accessory proteins bind to actin, controlling the number, length, position, and attachments of the actin filaments. Different cells and tissues contain different accessory proteins, which accounts for the different shapes and movements of different cells. For example, in some cells, actin filaments are bundled by accessory proteins, and the bundle is attached to the cell membrane to form microvilli, stable protrusions that resemble tiny bristles. Microvilli on the surface of epithelial cells such as those lining the intestine increase the cell’s surface area and thus facilitate the absorption of ingested food and water molecules. Other types of microvilli are involved in the detection of sound in the ear, where their movement, caused by sound waves, sends an electrical signal to the brain.
Many actin filaments in non-muscle cells have only a transient existence, polymerizing and depolymerizing in controlled ways that create movement. For example, many cells continually send out and retract tiny “filopodia,” long needlelike projections of the cell membrane that are thought to enable cells to probe their environment and decide which direction to go. Like microvilli, filopodia are formed when actin filaments push out the membrane, but, because these actin filaments are less stable, filopodia have only a brief existence. Another actin structure only transiently associated with the cell membrane is the contractile ring, which is composed of actin filaments running around the circumference of the cell during cell division. As its name implies, this ring pulls in the cell membrane by a myosin-dependent process, thereby pinching the cell in half.
Microtubules are long filaments formed from 13 to 15 protofilament strands of a globular subunit called tubulin, with the strands arranged in the form of a hollow cylinder. Like actin filaments, microtubules are polar, having “plus” and “minus” ends. Most microtubule plus ends are constantly growing and shrinking, by respectively adding and losing subunits at their ends. Stable microtubules are found in cilia and flagella. Cilia are hairlike structures found on the surface of certain types of epithelial cells, where they beat in unison to move fluid and particles over the cell surface. Cilia are closely related in structure to flagella. Flagella such as those found on sperm cells produce a helical wavelike motion that enables a cell to propel itself rapidly through fluids. In cilia and flagella a set of microtubules is connected in a regular array by numerous accessory proteins that act as links and spokes in the assembly. Movement of the cilia or flagella occurs when adjacent microtubules slide past one another, bending the structures. This motion is caused by the motor protein dynein, which uses the energy of ATP hydrolysis to move along the microtubules, in a manner resembling the movement of myosin along actin filaments.
In most cells, microtubules grow outward, from the cell centre to the cell membrane, from a special region of the cytoplasm near the nuclear envelope called the centrosome. The minus ends of these microtubules are embedded in the centrosome, while the plus ends terminate near the cell membrane. The plus ends grow and shrink rapidly, a process known as dynamic instability. At the start of cell division, the centrosome replicates and divides in two. The two centrosomes separate and move to opposite sides of the nuclear envelope, where each nucleates a starlike array of microtubules, forming the mitotic spindle. The mitotic spindle partitions the duplicated chromosomes into the two daughter cells during mitosis (see belowCell division and growth).
Microtubules often serve as tracks for the transport of membrane vesicles in the cell, carried by the motor proteins kinesin and dynein. Kinesins generally move toward the plus end of the microtubule, and dyneins move toward the minus end. Microtubule-based vesicle transport occurs in nearly all cells, but it is especially prominent in the long thin processes of neurons, carrying essential components to and from the synapses at the ends of the processes.
Intermediate filaments are so named because they are thicker than actin filaments and thinner than microtubules or muscle myosin filaments. The subunits of intermediate filaments are elongated, not globular, and are associated in an antipolar manner. As a result, the overall filament has no polarity, and therefore no motor proteins move along intermediate filaments. Intermediate filaments are found only in complex multicellular organisms. They are encoded by a large number of different genes and can be grouped into families based on their amino acid sequences. Cells in different tissues of the body express one or another of these genes at different times. One cell can even change which type of intermediate filament protein is expressed over its lifetime. Most likely, the different forms of intermediate filaments have subtle but critical differences in their functional characteristics, helping to define the function of the cell. In general, intermediate filaments serve as structural elements, helping cells maintain their shape and integrity. For example, keratin filaments, the intermediate filaments of epithelial cells, which line surfaces of the body, give strength to the cell sheet that covers the surface. Mutations in keratin genes can result in blisters when the epithelial cell sheet is weak and prone to rupture. Keratin mutations can also cause deformations in the hair, nails, and corneas. Another example of a family of intermediate filaments is the lamin family, which comprises the nuclear lamina, a fibrous shell that underlies and supports the nuclear membrane.
The Cell Matrix And Cell-To-Cell Communication
The Cell Matrix And Cell-To-Cell Communication
The Cell Matrix And Cell-To-Cell Communication (B)
The development of single cells into multicellular organisms involves a number of adaptations. The cells become specialized, acquiring distinct functions that contribute to the survival of the organism. The behaviour of individual cells is also integrated with that of similar cells, so that they act together in a regulated fashion. To achieve this integration, cells assemble into specialized tissues, each tissue being composed of cells and the spaces outside of the cells.
The surface of cells is important in coordinating their activities within tissues. Embedded in the plasma membrane of each cell are a number of proteins that interact with the surface or secretions of other cells. These proteins enable cells to “recognize” and adhere to the extracellular matrix and one another and to form populations distinct from surrounding cells. These interactions are key to the organizational behaviour of cell populations and contribute to the formation of embryonic tissues and the function of normal tissue in the adult organism.
Electron micrograph of a small area of dense fibrous connective tissue, illustrating the intimate association of cells and fibres. In the centre is a portion of a fibrocyte, and on either side are two collagen fibres. The collagen fibre on the left is cut transversely, showing round cross sections of the unit fibrils. The collagen fibre on the right has been cut nearly parallel to its long axis and shows extensive segments of the cross-striated fibrils. (Magnified about 6,625 ×.)
A substantial part of tissues is the space outside of the cells, called the extracellular space. This is filled with a composite material, known as the extracellular matrix, composed of a gel in which a number of fibrous proteins are suspended. The gel consists of large polysaccharide (complex sugar) molecules in a water solution of inorganic salts, nutrients, and waste products known as the interstitial fluid. The major types of protein in the matrix are structural proteins and adhesive proteins.
There are two general types of tissues distinct not only in their cellular organization but also in the composition of their extracellular matrix. The first type, mesenchymal tissue, is made up of clusters of cells grouped together but not closely adherent to one another. They synthesize a highly hydrated gel, rich in salts, fluid, and fibres, known as the interstitial matrix. Connective tissue is a mesenchyme that fastens together other more highly organized tissues. The solidity of various connective tissues varies according to the consistency of their extracellular matrix, which in turn depends on the water content of the gels, the amount and type of polysaccharides and structural proteins, and the presence of other salts. For example, bone is rich in calcium phosphate, giving that tissue its rigidity; tendons are mostly fibrous structural proteins, yielding a ropelike consistency; and joint spaces are filled with a lubricating fluid of mostly polysaccharide and interstitial fluid.
Epithelial tissues, the second type, are sheets of cells adhering at their side, or lateral, surfaces. They synthesize and deposit at their bottom, or basal, surfaces an organized complex of matrix materials known as the basal lamina or basement membrane. This thin layer serves as a boundary with connective tissue and as a substrate to which epithelial cells are attached.
The polysaccharides, or glycans, of the extracellular matrix are responsible for its gel-like quality and for organizing its components. These large acidic molecules exist alone (as glycosaminoglycans) or in combination with small proteins (as proteoglycans). They bind an extraordinarily large amount of water, thus forming massively swollen gels that fill the spaces between cells. Bound to proteins, they also organize other molecules in the extracellular matrix. The firmness and resiliency of cartilage, as at the surface of joints, is due to highly organized proteoglycans that bind water tightly.
Matrix proteins are large molecules tightly bound to form extensive networks of insoluble fibres. These fibres may even exceed the size of the cells themselves. The proteins are of two general types, structural and adhesive.
The structural proteins, collagen and elastin, are the dominant matrix proteins. At least 10 different types of collagen are present in various tissues. The most common, type I collagen, is the most abundant protein in vertebrate animals, accounting for nearly 25 percent of the total protein in the body. The various collagen types share structural features, all being composed of three intertwined polypeptide chains. In some collagens the chains are linked together by covalent bonds, yielding a ropelike structure of great tensile strength. Indeed, the toughness of leather, chemically treated animal skin, is due to its content of collagen. Elastin is also a cross-linked protein, but, instead of forming rigid coils, it imparts elasticity to tissues. Only one type of elastin is known; it varies in elasticity according to variations in its cross-linking.
The adhesive proteins of the extracellular matrix bind matrix molecules to one another and to cell surfaces. These proteins are modular in that they contain several functional domains packaged together in a single molecule. Each domain binds to a specific matrix component or to a specific site on a cell. The major adhesive protein of the interstitial matrix is called fibronectin; that of the basal lamina is known as laminin.
Molecules intimately associated with the cell membrane link cells to the extracellular matrix. These molecules, called matrix receptors, bind selectively to specific matrix components and interact, directly or indirectly, with actin protein fibres that form the cytoskeleton inside the cell. This association of actin fibres with matrix components via receptors on the cell membrane can influence the organization of membrane molecules as well as matrix components and can modify the shape and function of the cytoskeleton. Changes in the cytoskeleton can lead to changes in cell shape, movement, metabolism, and development.
The ability of cells to recognize and adhere to one another plays an important role in cell survival and reproduction. For example, when starved, several types of single-cell organisms band together to develop the specialized cells needed for reproduction. In this process, certain cells at the centre of the developing aggregate secrete chemicals that cause the other cells to adhere tightly into a group. In the case of slime moldamoebas, starvation causes the secretion of a compound, cyclic adenosine monophosphate (cyclic AMP, or CAMP), that induces the cells to stick together end to end. With further aggregation, the cells produce another cell-surface glycoprotein with which they stick to one another over their entire surfaces. The cellular aggregates then produce an extracellular matrix, which holds the cells together in a specific structural form.
Some multicellular animals or tissues can be dissociated into suspensions of single cells that show the same cellular recognition and adhesion as do aggregates of single-cell organisms. The marine sponge, for example, can be sieved through a mesh, yielding single cells and cells in clumps. When this cell suspension is rotated in culture, the cells reaggregate and in time reform a normal sponge. This reassociation shows selective cell recognition; that is, only cells of the same species reassociate. The ability of the cells to distinguish cells of their own species from those of others is mediated by proteoglycan molecules in the extracellular matrix. The proteoglycan binds to specific cell-surface receptor sites that are unique to a single species of sponge.
Multicellular animal.
Marine sponges are multicellular animals that can regenerate from single cells. The cells of a sponge rely on the processes of intercellular recognition and cellular adhesion to form aggregates of cells of the same species that eventually develop into an adult sponge.
Cells from tissues of vertebrate animals can, like sponge cells, be dissociated and allowed to reaggregate. For example, when vertebrate embryonic cells from two different tissues are dissociated and then rotated together in culture, the cells form a multicellular aggregate within which they sort according to the type of tissue, a sorting that occurs regardless of whether the cells are from the same or different species. The specificity is due to a set of cell-surface glycoproteins called cell adhesion molecules (CAM). A portion of the CAM that extends from the surface of a cell adheres to identical molecules on the surface of adjacent cells. These CAM appear early in embryonic life, and their amounts in tissues change as the organs develop. The CAM, however, are not responsible for the stable adhesion of one cell to another; this more permanent adhesion is carried out by cell junctions.
There are three functional categories of cell junction: adhering junctions, often called desmosomes; tight, or occluding, junctions; and gap, or permeable, junctions. Adhering junctions hold cells together mechanically and are associated with intracellular fibres of the cytoskeleton. Tight junctions also hold cells together, but they form a nearly leakproof intercellular seal by fusion of adjacent cell membranes. Both adhering junctions and tight junctions are present primarily in epithelial cells. Many cell types also possess gap junctions, which allow small molecules to pass from one cell to the next through a channel.
Adhering junctions
Cells subject to abrasion or other mechanical stress, such as those of the surface epithelia of the skin, have junctions that adhere cells to one another and to the extracellular matrix. These adhering junctions are called desmosomes when occurring between cells and hemidesmosomes (half-desmosomes) when linked to the matrix. Adhering junctions distribute mechanical shear force throughout the tissue and to the underlying matrix by virtue of their association with intermediate filaments crossing the interior of the cell. The linkage of these filaments, also called keratin filaments, to the desmosomes and, through these junctions, to adjacent cells provides a nearly continuous fibrous network throughout an epithelial sheet. Adhering junctions are also seen in other types of cells—for example, in the muscles of the heart and uterus—allowing these cells to remain anchored together despite the contractions of the muscles.
Tight junctions
Sheets of cells separate fluids within the organs from fluids outside, as in the epithelial layer lining the intestine. This separation requires leakproof junctions between cells. Tight junctions form leakproof seals by fusing the plasma membranes of adjacent cells, creating a continuous barrier through which molecules cannot pass. The membranes are fused by tight associations of two types of specialized integral membrane proteins, in turn repelling large water-soluble molecules. In invertebrates this function is provided by septate junctions, in which the proteins of the membrane rather than the lipids form the seal.
These junctions allow communication between adjacent cells via the passage of small molecules directly from the cytoplasm of one cell to that of another. Molecules that can pass between cells coupled by gap junctions include inorganic salts, sugars, amino acids,nucleotides, and vitamins but not large molecules such as proteins or nucleic acids.
Gap junctions are crucial to the integration of certain cellular activities. For example, heart muscle cells generate electrical current by the movement of inorganic salts. If the cells are coupled, they will share this electrical current, allowing the synchronous contraction of all the cells in the tissue. This coupling function requires the regulation of molecular traffic through the gaps. The junctions are not open pores but dynamic channels, which change their permeability with changes in cellular activity. They consist of proteins completely crossing the cell membrane as six-sided columns with central pores. Under certain conditions the proteins are thought to change shape, causing the pores to become smaller or larger and thus changing the permeability of the junction.
Gap junctions are also found in tissues that are not electrically active. In these tissues, the junctions allow nutrients and waste products to travel throughout the tissue. Cells in such tissues are said to be metabolically coupled. During the formation of embryos, gap junctions are crucial to establishing differences between separate groups of cells, the coupled cells undergoing development together to become a specialized tissue.
Cell-to-cell communication via chemical signaling
Cell-to-cell communication via chemical signaling (B)
In addition to cell-matrix and cell-cell interactions, cell behaviour in multicellular organisms is coordinated by the passage of chemical or electrical signals between cells. The most common form of chemical signaling is via molecules secreted from the cells and moving through the extracellular space. Signaling molecules may also remain on cell surfaces, influencing other cells only after the cells make physical contact. Finally, as noted above, gap junctions allow small molecules to move between the cytoplasms of adjacent cells.
Chemical signals secreted by cells can act over varying distances. In the autocrine signaling process, molecules act on the same cells that produce them. In paracrine signaling, they act on nearby cells. Autocrine signals include extracellular matrix molecules and various factors that stimulate cell growth. An example of paracrine signals is the chemical transmitted from nerve to muscle that causes the muscle to contract. In this instance, the muscle cells have regions specialized to receive chemical signals from an adjacent nerve cell. In both autocrine and paracrine signaling, the chemical signal works in the immediate vicinity of the cell that produces it and is present at high concentrations. A chemical signal picked up by the bloodstream and taken to distant sites is called an endocrine signal. Most hormones produced in vertebrates are endocrine signals, such as the hormones produced in the pituitary gland at the base of the brain and carried by the bloodstream to act at low concentrations on the thyroid or adrenal glands.
The concentration at which a chemical signal acts has significance for its target cell. Chemical signals that act at high concentration act locally and rapidly. On the other hand, chemical signals that act at low concentrations act at distances and are generally slow.
Hormones and active metabolites bind to different types of receptors. Water-soluble molecules (i.e., insulin) cannot pass through the lipid membrane of a cell and thus rely on cell surface receptors to transmit messages to the interior of the cell. In contrast, lipid-soluble molecules (i.e., certain active metabolites) are able to diffuse through the lipid membrane to communicate messages directly to the nucleus.
The ability of a cell to respond to an extracellular signal depends on the presence of specific proteins called receptors, which are located on the cell surface or in the cytoplasm. Receptors bind chemical signals that ultimately trigger a mechanism to modify the behaviour of the target cell. Cells may contain an array of specific receptors that allow them to respond to a variety of chemical signals.
Signal molecules are either soluble or insoluble. Water-soluble molecules, such as the polypeptide hormoneinsulin, bind to receptors at cell surfaces. On the other hand, lipid-soluble molecules, such as the steroid hormones produced by the ovary or testis, pass through the lipid bilayer of the cell membrane to reach receptors within the cytoplasm. Extracellular matrix molecules are chemical signals, but, because of their size and insolubility, they act only on cell surface receptors and are neither taken up by the cells nor rapidly destroyed.
The binding of chemical signals to their corresponding receptors induces events within the cell that ultimately change its behaviour. The nature of these intracellular events differs according to the type of receptor. Also, the same chemical signal can trigger different responses in different types of cell.
Cell surface receptors work in several ways when they are occupied. Some receptors enter the cell still bound to the chemical signal. Others activate membraneenzymes, which produce certain intracellular chemical mediators. Still other receptors open membrane channels, allowing a flow of ions that causes either a change in the electrical properties of the membrane or a change in the ion concentration in the cytoplasm. This regulation of enzymes or membrane channels produces changes in the concentration of intracellular signaling molecules, which are often called second messengers (the first messenger being the extracellular chemical signal bound to the receptor).
Two common intracellular signaling molecules are cyclic AMP and the calcium ion. Cyclic AMP is a derivative of adenosine triphosphate, the ubiquitous energy-carrying molecule of the cell. The intracellular concentrations of both cyclic AMP and calcium ions are normally very low. The binding of an extracellular chemical signal to a cell surface receptor stimulates an enzyme complex in the membrane to produce cyclic AMP. This second messenger then diffuses into the cytoplasm and acts on intracellular enzymes called kinases that modify the behaviour of the cell, culminating in the activation of target genes that increase the synthesis of certain proteins. The action of cyclic AMP is brief because it is rapidly degraded by specific enzymes.
Occupancy of other surface receptors causes a transient opening of membrane channels. This can allow calcium ions to enter the cytoplasm from the extracellular space, where their concentration is higher. The action of calcium ions is also brief because they are rapidly pumped out of the cell or bound to intracellular molecules, lowering the cytoplasmic concentration to the state existing before stimulation.
Some extracellular chemical signals enter the cell intact, still bound to the receptor, without generating a second messenger. In this mechanism, receptor occupancy causes individual receptors within the cell membrane to aggregate spontaneously. That portion of the membrane containing the aggregated receptors is then taken into the cell, where it fuses with various membrane-bounded organelles in the cytoplasm. In some instances the chemical signal is released within the organelles, and in almost all instances the ingested membrane is rapidly returned to the cell membrane along with the surface receptors.
The plantcell wall is a specialized form of extracellular matrix that surrounds every cell of a plant and is responsible for many of the characteristics distinguishing plant from animal cells. Although often perceived as an inactive product serving mainly mechanical and structural purposes, the cell wall actually has a multitude of functions upon which plant life depends. Such functions include: (1) providing the protoplast, or living cell, with mechanical protection and a chemically buffered environment, (2) providing a porous medium for the circulation and distribution of water, minerals, and other small nutrient molecules, (3) providing rigid building blocks from which stable structures of higher order, such as leaves and stems, can be produced, and (4) providing a storage site of regulatory molecules that sense the presence of pathogenic microbes and control the development of tissues.
All cell walls contain two layers, the middle lamella and the primary cell wall, and many cells produce an additional layer, called the secondary wall. The middle lamella serves as a cementing layer between the primary walls of adjacent cells. The primary wall is the cellulose-containing layer laid down by cells that are dividing and growing. To allow for cell wall expansion during growth, primary walls are thinner and less rigid than those of cells that have stopped growing. A fully grown plant cell may retain its primary cell wall (sometimes thickening it), or it may deposit an additional, rigidifying layer of different composition; this is the secondary wall. Secondary cell walls are responsible for most of the plant’s mechanical support as well as the mechanical properties prized in wood. In contrast to the permanent stiffness and load-bearing capacity of thick secondary walls, the thin primary walls are capable of serving a structural, supportive role only when the vacuoles within the cell are filled with water to the point that they exert a turgor pressure against the cell wall. Turgor-induced stiffening of primary walls is analogous to the stiffening of the sides of a pneumatic tire by air pressure. The wilting of flowers and leaves is caused by a loss of turgor pressure, which results in turn from the loss of water from the plant cells.
Although primary and secondary wall layers differ in detailed chemical composition and structural organization, their basic architecture is the same, consisting of cellulose fibres of great tensile strength embedded in a water-saturated matrix of polysaccharides and structural glycoproteins.
Cellulose
Cellulose consists of several thousand glucose molecules linked end to end. The chemical links between the individual glucose subunits give each cellulose molecule a flat ribbonlike structure that allows adjacent molecules to band laterally together into microfibrils with lengths ranging from two to seven micrometres. Cellulose fibrils are synthesized by enzymes floating in the cell membrane and are arranged in a rosette configuration. Each rosette appears capable of “spinning” a microfibril into the cell wall. During this process, as new glucose subunits are added to the growing end of the fibril, the rosette is pushed around the cell on the surface of the cell membrane, and its cellulose fibril becomes wrapped around the protoplast. Thus, each plant cell can be viewed as making its own cellulose fibril cocoon.
Matrix polysaccharides
The two major classes of cell wall matrix polysaccharides are the hemicelluloses and the pectic polysaccharides, or pectins. Both are synthesized in the Golgi apparatus, brought to the cell surface in small vesicles, and secreted into the cell wall.
Hemicelluloses consist of glucose molecules arranged end to end as in cellulose, with short side chains of xylose and other uncharged sugars attached to one side of the ribbon. The other side of the ribbon binds tightly to the surface of cellulose fibrils, thereby coating the microfibrils with hemicellulose and preventing them from adhering together in an uncontrolled manner. Hemicellulose molecules have been shown to regulate the rate at which primary cell walls expand during growth.
The heterogeneous, branched, and highly hydrated pectic polysaccharides differ from hemicelluloses in important respects. Most notably, they are negatively charged because of galacturonic acid residues, which, together with rhamnose sugar molecules, form the linear backbone of all pectic polysaccharides. The backbone contains stretches of pure galacturonic acid residues interrupted by segments in which galacturonic acid and rhamnose residues alternate; attached to these latter segments are complex, branched sugar side chains. Because of their negative charge, pectic polysaccharides bind tightly to positively charged ions, or cations. In cell walls, calcium ions cross-link the stretches of pure galacturonic acid residues tightly, while leaving the rhamnose-containing segments in a more open, porous configuration. This cross-linking creates the semirigid gel properties characteristic of the cell wall matrix—a process exploited in the preparation of jellied preserves.
Proteins
Although plant cell walls contain only small amounts of protein, they serve a number of important functions. The most prominent group are the hydroxyproline-rich glycoproteins, shaped like rods with connector sites, of which extensin is a prominent example. Extensin contains 45 percent hydroxyproline and 14 percent serine residues distributed along its length. Every hydroxyproline residue carries a short side chain of arabinose sugars, and most serine residues carry a galactose sugar; this gives rise to long molecules, resembling bottle brushes, that are secreted into the cell wall toward the end of primary-wall formation and become covalently cross-linked into a mesh at the time that cell growth stops. Plant cells may control their ultimate size by regulating the time at which this cross-linking of extensin molecules occurs.
In addition to the structural proteins, cell walls contain a variety of enzymes. Most notable are those that cross-link extensin, lignin, cutin, and suberin molecules into networks. Other enzymes help protect plants against fungal pathogens by breaking fragments off of the cell walls of the fungi. The fragments in turn induce defense responses in underlying cells. The softening of ripe fruit and dropping of leaves in the autumn are brought about by cell wall-degrading enzymes.
Plastics
Cell wall plastics such as lignin,cutin, and suberin all contain a variety of organic compounds cross-linked into tight three-dimensional networks that strengthen cell walls and make them more resistant to fungal and bacterial attack. Lignin is the general name for a diverse group of polymers of aromatic alcohols. Deposited mostly in secondary cell walls and providing the rigidity of terrestrial vascular plants, it accounts for up to 30 percent of a plant’s dry weight. The diversity of cross-links between the polymers—and the resulting tightness—makes lignin a formidable barrier to the penetration of most microbes.
Cutin and suberin are complex biopolyesters composed of fatty acids and aromatic compounds. Cutin is the major component of the cuticle, the waxy, water-repelling surface layer of cell walls exposed to the environment aboveground. By reducing the wetability of leaves and stems—and thereby affecting the ability of fungal spores to germinate—it plays an important part in the defense strategy of plants. Suberin serves with waxes as a surface barrier of underground parts. Its synthesis is also stimulated in cells close to wounds, thereby sealing off the wound surfaces and protecting underlying cells from dehydration.
Similar to the gap junction of animal cells is the plasmodesma, a channel passing through the cell wall and allowing direct molecular communication between adjacent plant cells. Plasmodesmata are lined with cell membrane, in effect uniting all connected cells with one continuous cell membrane. Running down the middle of each channel is a thin membranous tube that connects the endoplasmic reticula (ER) of the two cells. This structure is a remnant of the ER of the original parent cell, which, as the parent cell divided, was caught in the developing cell plate.
Although the precise mechanisms are not fully understood, the plasmodesma is thought to regulate the passage of small molecules such as salts, sugars, and amino acids by constricting or dilating the openings at each end of the channel.
Oligosaccharides with regulatory functions
The discovery of cell wall fragments with regulatory functions opened a new era in plant research. For years scientists had been puzzled by the chemical complexity of cell wall polysaccharides, which far exceeds the structural requirements of plant cell walls. The answer came when it was found that specific fragments of cell wall polysaccharides, called oligosaccharins, are able to induce specific responses in plant cells and tissues. One such fragment, released by enzymes used by fungi to break down plant cell walls, consists of a linear polymer of 10 to 12 galacturonic acid residues. Exposure of plant cells to such fragments induces them to produce antibiotics known as phytoalexins. In other experiments it has been shown that exposing strips of tobacco stem cells to a different type of cell wall fragment leads to the growth of roots; other fragments lead to the formation of stems, and yet others to the production of flowers. In all instances the concentration of oligosaccharins required to bring about the observed responses is equal to that of hormones in animal cells; indeed, oligosaccharins may be viewed as the oligosaccharide hormones of plants.
Cell Division And Growth
Cell Division And Growth
Cell Division And Growth (B)
In unicellular organisms, cell division is the means of reproduction; in multicellular organisms, it is the means of tissuegrowth and maintenance. Survival of the eukaryotes depends upon interactions between many cell types, and it is essential that a balanced distribution of types be maintained. This is achieved by the highly regulated process of cell proliferation. The growth and division of different cell populations are regulated in different ways, but the basic mechanisms are similar throughout multicellular organisms.
Mitosis.
One cell gives rise to two genetically identical daughter cells during the process of mitosis.
Most tissues of the body grow by increasing their cell number, but this growth is highly regulated to maintain a balance between different tissues. In adults most cell division is involved in tissue renewal rather than growth, many types of cells undergoing continuous replacement. Skin cells, for example, are constantly being sloughed off and replaced; in this case, the mature differentiated cells do not divide, but their population is renewed by division of immature stem cells. In certain other cells, such as those of the liver, mature cells remain capable of division to allow growth or regeneration after injury.
In contrast to these patterns, other types of cells either cannot divide or are prevented from dividing by certain molecules produced by nearby cells. As a result, in the adult organism, some tissues have a greatly reduced capacity to renew damaged or diseased cells. Examples of such tissues include heart muscle, nerve cells of the central nervous system, and lens cells in mammals. Maintenance and repair of these cells is limited to replacing intracellular components rather than replacing entire cells.
Before a cell can divide, it must accurately and completely duplicate the genetic information encoded in its DNA in order for its progeny cells to function and survive. This is a complex problem because of the great length of DNA molecules. Each humanchromosome consists of a long double spiral, or helix, each strand of which consists of more than 100 million nucleotides (see aboveThe nucleus).
The duplication of DNA is called DNA replication, and it is initiated by complex enzymes called DNA polymerases. These progress along the molecule, reading the sequences of nucleotides that are linked together to make DNA chains. Each strand of the DNA double helix, therefore, acts as a template specifying the nucleotide structure of a new growing chain. After replication, each of the two daughter DNA double helices consists of one parental DNA strand wound around one newly synthesized DNA strand.
In order for DNA to replicate, the two strands must be unwound from each other. Enzymes called helicases unwind the two DNA strands, and additional proteins bind to the separated strands to stabilize them and prevent them from pairing again. In addition, a remarkable class of enzyme called DNA topoisomerase removes the helical twists by cutting either one or both strands and then resealing the cut. These enzymes can also untangle and unknot DNA when it is tightly coiled into a chromatin fibre.
In the circular DNA of prokaryotes, replication starts at a unique site called the origin of replication and then proceeds in both directions around the molecule until the two processes meet, producing two daughter molecules. In rapidly growing prokaryotes, a second round of replication can start before the first has finished. The situation in eukaryotes is more complicated, as replication moves more slowly than in prokaryotes. At 500 to 5,000 nucleotides per minute (versus 100,000 nucleotides per minute in prokaryotes), it would take a human chromosome about a month to replicate if started at a single site. Actually, replication begins at many sites on the long chromosomes of animals,plants, and fungi. Distances between adjacent initiation sites are not always the same; for example, they are closer in the rapidly dividing embryonic cells of frogs or flies than in adult cells of the same species.
Accurate DNA replication is crucial to ensure that daughter cells have exact copies of the genetic information for synthesizing proteins. Accuracy is achieved by a “proofreading” ability of the DNA polymerase itself. It can erase its own errors and then synthesize anew. There are also repair systems that correct genetic damage to DNA. For example, the incorporation of an incorrect nucleotide, or damage caused by mutagenic agents, can be corrected by cutting out a section of the daughter strand and recopying the parental strand.
In eukaryotes the processes of DNA replication and cell division occur at different times of the cell division cycle. During cell division, DNA condenses to form short, tightly coiled, rodlike chromosomes. Each chromosome then splits longitudinally, forming two identical chromatids. Each pair of chromatids is divided between the two daughter cells during mitosis, or division of the nucleus, a process in which the chromosomes are propelled by attachment to a bundle of microtubules called the mitotic spindle.
Mitosis can be divided into five phases. In prophase the mitotic spindle forms and the chromosomes condense. In prometaphase the nuclear envelope breaks down (in many but not all eukaryotes) and the chromosomes attach to the mitotic spindle. Both chromatids of each chromosome attach to the spindle at a specialized chromosomal region called the kinetochore. In metaphase the condensed chromosomes align in a plane across the equator of the mitotic spindle. Anaphase follows as the separated chromatids move abruptly toward opposite spindle poles. Finally, in telophase a new nuclear envelope forms around each set of unraveling chromatids.
An essential feature of mitosis is the attachment of the chromatids to opposite poles of the mitotic spindle. This ensures that each of the daughter cells will receive a complete set of chromosomes. The mitotic spindle is composed of microtubules, each of which is a tubular assembly of molecules of the protein tubulin (see aboveThe cytoskeleton). Some microtubules extend from one spindle pole to the other, while a second class extends from one spindle pole to a chromatid. Microtubules can grow or shrink by the addition or removal of tubulin molecules. The shortening of spindle microtubules at anaphase propels attached chromatids to the spindle poles, where they unravel to form new nuclei.
The two poles of the mitotic spindle are occupied by centrosomes, which organize the microtubule arrays. In animal cells each centrosome contains a pair of cylindrical centrioles, which are themselves composed of complex arrays of microtubules. Centrioles duplicate at a precise time in the cell division cycle, usually close to the start of DNA replication.
After mitosis comes cytokinesis, the division of the cytoplasm. This is another process in which animal and plant cells differ. In animal cells cytokinesis is achieved through the constriction of the cell by a ring of contractile microfilaments consisting of actin and myosin, the proteins involved in muscle contraction and other forms of cell movement. In plant cells the cytoplasm is divided by the formation of a new cell wall, called the cell plate, between the two daughter cells. The cell plate arises from small Golgi-derived vesicles that coalesce in a plane across the equator of the late telophase spindle to form a disk-shaped structure. In this process, each vesicle contributes its membrane to the forming cell membranes and its matrix contents to the forming cell wall. A second set of vesicles extends the edge of the cell plate until it reaches and fuses with the sides of the parent cell, thereby completely separating the two new daughter cells. At this point, cellulose synthesis commences, and the cell plate becomes a primary cell wall (see aboveThe plant cell wall).
A specialized division of chromosomes called meiosis occurs during the formation of the reproductive cells, or gametes, of sexually reproducing organisms. Gametes such as ova,sperm, and pollen begin as germ cells, which, like other types of cells, have two copies of each gene in their nuclei. The chromosomes composed of these matching genes are called homologs. During DNA replication, each chromosome duplicates into two attached chromatids. The homologous chromosomes are then separated to opposite poles of the meiotic spindle by microtubules similar to those of the mitotic spindle. At this stage in the meiosis of germ cells, there is a crucial difference from the mitosis of other cells. In meiosis the two chromatids making up each chromosome remain together, so that whole chromosomes are separated from their homologous partners. Cell division then occurs, followed by a second division that resembles mitosis more closely in that it separates the two chromatids of each remaining chromosome. In this way, when meiosis is complete, each mature gamete receives only one copy of each gene instead of the two copies present in other cells.
Gamete; meiosis.
The formation of gametes (sex cells) occurs during the process of meiosis.
Meiosis in pollen.
Cross section of a plant anther showing meiosis occurring in pollen grain cells.
In prokaryotes,DNA synthesis can take place uninterrupted between cell divisions, and new cycles of DNA synthesis can begin before previous cycles have finished. In contrast, eukaryotes duplicate their DNA exactly once during a discrete period between cell divisions. This period is called the S (for synthetic) phase. It is preceded by a period called G1 (meaning “first gap”) and followed by a period called G2, during which nuclear DNA synthesis does not occur.
The four periods G1, S, G2, and M (for mitosis) make up the cell division cycle. The cell cycle characteristically lasts between 10 and 20 hours in rapidly proliferating adult cells, but it can be arrested for weeks or months in quiescent cells or for a lifetime in neurons of the brain. Prolonged arrest of this type usually occurs during the G1 phase and is sometimes referred to as G0. In contrast, some embryonic cells, such as those of fruit flies (vinegar flies), can complete entire cycles and divide in only 11 minutes. In these exceptional cases, G1 and G2 are undetectable, and mitosis alternates with DNA synthesis. In addition, the duration of the S phase varies dramatically. The fruit flyembryo takes only four minutes to replicate its DNA, compared with several hours in adult cells of the same species.
Controlled proliferation
Controlled proliferation (B)
Several studies have identified the transition from the G1 to the S phase as a crucial control point of the cell cycle. Stimuli are known to cause resting cells to proliferate by inducing them to leave G1 and begin DNA synthesis. These stimuli, called growth factors, are naturally occurring proteins specific to certain groups of cells in the body. They include nerve growth factor, epidermal growth factor, and platelet-derived growth factor. Such factors may have important roles in the healing of wounds as well as in the maintenance and growth of normal tissues. Many growth factors are known to act on the external membrane of the cell, by interacting with specialized proteinreceptor molecules. These respond by triggering further cellular changes, including an increase in calcium levels that makes the cell interior more alkaline and the addition of phosphate groups to the amino acidtyrosine in proteins. The complex response of cells to growth factors is of fundamental importance to the control of cell proliferation.
Cancer can arise when the controlling factors over cell growth fail and allow a cell and its descendants to keep dividing at the expense of the organism. Studies of viruses that transform cultured cells and thus lead to the loss of control of cell growth have provided insight into the mechanisms that drive the formation of tumours. Transformed cells may differ from their normal progenitors by continuing to proliferate at very high densities, in the absence of growth factors, or in the absence of a solid substrate for support.
Cancer-causing retroviruses.
Retroviral insertion can convert a proto-oncogene, integral to the control of cell division, into an oncogene, the agent responsible for transforming a healthy cell into a cancer cell. An acutely transforming retrovirus (shown at top), which produces tumours within weeks of infection, incorporates genetic material from a host cell into its own genome upon infection, forming a viral oncogene. When the viral oncogene infects another cell, an enzyme called reverse transcriptase copies the single-stranded genetic material into double-stranded DNA, which is then integrated into the cellular genome. A slowly transforming retrovirus (shown at bottom), which requires months to elicit tumour growth, does not disrupt cellular function through the insertion of a viral oncogene. Rather, it carries a promoter gene that is integrated into the cellular genome of the host cell next to or within a proto-oncogene, allowing conversion of the proto-oncogene to an oncogene.
Major advances in the understanding of growth control have come from studies of the viral genes that cause transformation. These viral oncogenes have led to the identification of related cellular genes called protooncogenes. Protooncogenes can be altered by mutation or epigenetic modification, which converts them into oncogenes and leads to cell transformation. Specific oncogenes are activated in particular human cancers. For example, an oncogene called RAS is associated with many epithelial cancers, while another, called MYC, is associated with leukemias.
An interesting feature of oncogenes is that they may act at different levels corresponding to the multiple steps seen in the development of cancer. Some oncogenes immortalize cells so that they divide indefinitely, whereas normal cells die after a limited number of generations. Other oncogenes transform cells so that they grow in the absence of growth factors. A combination of these two functions leads to loss of proliferation control, whereas each of these functions on its own cannot. The mode of action of oncogenes also provides important clues to the nature of growth control and cancer. For example, some oncogenes are known to encode receptors for growth factors that may cause continuous proliferation in the absence of appropriate growth factors.
Loss of growth control has the added consequence that cells no longer repair their DNA effectively, and thus aberrant mitoses occur. As a result, additional mutations arise that subvert a cell’s normal constraints to remain in its tissue of origin. Epithelial tumour cells, for example, acquire the ability to cross the basal lamina and enter the bloodstream or lymphatic system, where they migrate to other parts of the body, a process called metastasis. When cells metastasize to distant tissues, the tumour is described as malignant, whereas prior to metastasis a tumour is described as benign.
Adult organisms are composed of a number of distinct cell types. Cells are organized into tissues, each of which typically contains a small number of cell types and is devoted to a specific physiological function. For example, the epithelial tissue lining the small intestine contains columnar absorptive cells, mucus-secreting goblet cells, hormone-secreting endocrine cells, and enzyme-secreting Paneth cells. In addition, there exist undifferentiated dividing cells that lie in the crypts between the intestinal villi and serve to replace the other cell types when they become damaged or worn out. Another example of a differentiated tissue is the skeletal tissue of a long bone, which contains osteoblasts (large cells that synthesize bone) in the outer sheath and osteocytes (mature bone cells) and osteoclasts (multinucleate cells involved in bone remodeling) within the matrix.
Small intestine.
The small intestine contains many distinct types of cells, each of which serves a specific function.
Leaf structure. Structures of a leaf. The epidermis is often covered with a waxy protective cuticle that helps prevent water loss from inside the leaf. Oxygen, carbon dioxide, and water enter and exit the leaf through pores (stomata) scattered mostly along the lower epidermis. The stomata are opened and closed by the contraction and expansion of surrounding guard cells. The vascular, or conducting, tissues are known as xylem and phloem; water and minerals travel up to the leaves from the roots through the xylem, and sugars made by photosynthesis are transported to other parts of the plant through the phloem. Photosynthesis occurs within the chloroplast-containing mesophyll layer.
In general, the simpler the overall organization of the animal, the fewer the number of distinct cell types that they possess. Mammals contain more than 200 different cell types, whereas simple invertebrate animals may have only a few different types. Plants are also made up of differentiated cells, but they are quite different from the cells of animals. For example, a leaf in a higher plant is covered with a cuticle layer of epidermal cells. Among these are pores composed of two specialized cells, which regulate gaseous exchange across the epidermis. Within the leaf is the mesophyll, a spongy tissue responsible for photosynthetic activity. There are also veins composed of xylem elements, which transport water up from the soil, and phloem elements, which transport products of photosynthesis to the storage organs.
The various cell types have traditionally been recognized and classified according to their appearance in the light microscope following the process of fixing, processing, sectioning, and staining tissues that is known as histology. Classical histology has been augmented by a variety of more discriminating techniques. Electron microscopy allows for higher magnifications. Histochemistry involves the use of coloured precipitating substrates to stain particular enzymes in situ. Immunohistochemistry uses specific antibodies to identify particular substances, usually proteins or carbohydrates, within cells. In situ hybridization involves the use of nucleic acid probes to visualize the location of specific messenger RNAs (mRNA). These modern methods have allowed the identification of more cell types than could be visualized by classical histology, particularly in the brain, the immune system, and among the hormone-secreting cells of the endocrine system.
The biochemical basis of cell differentiation is the synthesis by the cell of a particular set of proteins, carbohydrates, and lipids. This synthesis is catalyzed by proteins called enzymes. Each enzyme in turn is synthesized in accordance with a particular gene, or sequence of nucleotides in the DNA of the cell nucleus. A particular state of differentiation, then, corresponds to the set of genes that is expressed and the level to which it is expressed.
Dolly the sheep; cloning.
Dolly the sheep was successfully cloned in 1996 by fusing the nucleus from a mammary-gland cell of a Finn Dorset ewe into an enucleated egg cell taken from a Scottish Blackface ewe. Carried to term in the womb of another Scottish Blackface ewe, Dolly was a genetic copy of the Finn Dorset ewe.
It is believed that all of an organism’s genes are present in each cell nucleus, no matter what the cell type, and that differences between tissues are not due to the presence or absence of certain genes but are due to the expression of some and the repression of others. In animals the best evidence for retention of the entire set of genes comes from whole animal cloning experiments in which the nucleus of a differentiated cell is substituted for the nucleus of a fertilized egg. In many species this can result in the development of a normal embryo that contains the full range of body parts and cell types. Likewise, in plants it is often possible to grow complete embryos from individual cells in tissue culture. Such experiments show that any nucleus has the genetic information required for the growth of a developing organism, and they strongly suggest that, for most tissues, cell differentiation arises from the regulation of genetic activity rather than the removal or destruction of unwanted genes. The only known exception to this rule comes from the immune system, where segments of DNA in developing white blood cells are slightly rearranged, producing a wide variety of antibody and receptor molecules. (See aboveRearrangement and modification of DNA.)
At the molecular level there are many ways in which the expression of a gene can be differentially regulated in different cell types. There may be differences in the copying, or transcription, of the gene into RNA; in the processing of the initial RNA transcript into mRNA; in the control of mRNA movement to the cytoplasm; in the translation of mRNA to protein; or in the stability of mRNA. However, the control of transcription has the most influence over gene expression and has received the most detailed analysis.
The DNA in the cell nucleus exists in the form of chromatin, which is made up of DNA bound to histones (simple alkaline proteins) and other nonhistone proteins. Most of the DNA is complexed into repeating structures called nucleosomes, each of which contains eight molecules of histone. Active genes are found in parts of the DNA where the chromatin has an “open” configuration, in which regulatory proteins are able to gain access to the DNA. The degree to which the chromatin opens depends on chemical modifications of the outer parts of the histone molecules and on the presence or absence of particular nonhistone proteins. Transcriptional control is exerted with the help of regulatory sequences that are found associated with a gene, such as the promoter sequence, a region near the start of the gene, and enhancer sequences, regions that lie elsewhere within the DNA that augment the activity of enzymes involved in the process of transcription. Whether or not transcription occurs depends on the binding of transcription factors to these regulatory sequences. Transcription factors are proteins that usually possess a DNA-binding region, which recognizes the specific regulatory sequence in the DNA, and an effector region, which activates or inhibits transcription. Transcription factors often work by recruiting enzymes that add modifications (e.g., acetyl groups or methyl groups) to or remove modifications from the outer parts of the histone molecules. This controls the folding of the chromatin and the accessibility of the DNA to RNA polymerase and other transcription factors.
In general, it requires several transcription factors working in combination to activate a gene. For example, the chicken delta 1 crystallin gene, normally expressed only in the lens of the eye, has a promoter that contains binding sites for two activating transcription factors and an enhancer that contains binding sites for two other activating transcription factors. There is also an additional enhancer site that can bind either an activator (deltaEF3) or a repressor (deltaEF1). Successful transcription requires that all these sites are occupied by the correct transcription factors.
Fully differentiated cells are qualitatively different from one another. States of terminal differentiation are stable and persistent, both in the lifetime of the cell and in successive cell generations (in the case of differentiated types that are capable of continued cell division). The inherent stability of the differentiated state is maintained by various processes, including feedback activation of genes by their own products and repression of inactive genes. Chromatin structure may be important in maintaining states of differentiation, although it is still unclear whether this can be maintained during DNA replication, which involves temporary removal of chromosomal proteins and unwinding of the DNA double helix.
A type of differentiation control that is maintained during DNA replication is the methylation of DNA, which tends to recruit histone deacetylases and hence close up the structure of the chromatin. DNA methylation occurs when a methyl group is attached to the exterior, or sugar-phosphate side, of a cytosine (C) residue. Cytosine methylation occurs only on a C nucleotide when it is connected to a G (guanine) nucleotide on the same strand of DNA. These nucleotide pairings are called CG dinucleotides. One class of DNA methylase enzyme can introduce new methylations when required, whereas another class, called maintenance methylases, methylates CG dinucleotides in the DNA double helix only when the CG of the complementary strand is already methylated. Each time the methylated DNA is replicated, the old strand has the methyl groups and the new strand does not. The maintenance methylase will then add methyl groups to all the CGs opposite the existing methyl groups to restore a fully methylated double helix. This mechanism guarantees stability of the DNA methylation pattern, and hence the differentiated state, during the processes of DNA replication and cell division.
Differentiation from visibly undifferentiated precursor cells occurs during embryonicdevelopment, during metamorphosis of larval forms, and following the separation of parts in asexual reproduction. It also takes place in adult organisms during the renewal of tissues and the regeneration of missing parts. Thus, cell differentiation is an essential and ongoing process at all stages of life.
The visible differentiation of cells is only the last of a progressive sequence of states. In each state, the cell becomes increasingly committed toward one type of cell into which it can develop. States of commitment are sometimes described as “specification” to represent a reversible type of commitment and as “determination” to represent an irreversible commitment. Although states of specification and determination both represent differential gene activity, the properties of embryonic cells are not necessarily the same as those of fully differentiated cells. In particular, cells in specification states are usually not stable over prolonged periods of time.
Two mechanisms bring about altered commitments in the different regions of the early embryo: cytoplasmic localization and induction. Cytoplasmic localization is evident in the earliest stages of development of the embryo. During this time, the embryo divides without growth, undergoing cleavage divisions that produce separate cells called blastomeres. Each blastomere inherits a certain region of the original eggcytoplasm, which may contain one or more regulatory substances called cytoplasmic determinants. When the embryo has become a solid mass of blastomeres (called a morula), it generally consists of two or more differently committed cell populations—a result of the blastomeres having incorporated different cytoplasmic determinants. Cytoplasmic determinants may consist of mRNA or protein in a particular state of activation. An example of the influence of a cytoplasmic determinant is a receptor called Toll, located in the membranes of Drosophila (fruit fly) eggs. Activation of Toll ensures that the blastomeres will develop into ventral (underside) structures, while blastomeres containing inactive Toll will produce cells that will develop into dorsal (back) structures.
In induction, the second mechanism of commitment, a substance secreted by one group of cells alters the development of another group. In early development, induction is usually instructive; that is, the tissue assumes a different state of commitment in the presence of the signal than it would in the absence of the signal. Inductive signals often take the form of concentration gradients of substances that evoke a number of different responses at different concentrations. This leads to the formation of a sequence of groups of cells, each in a different state of specification. For example, in Xenopus (clawed frog) the early embryo contains a signaling centre called the organizer that secretes inhibitors of bone morphogenetic proteins (BMPs), leading to a ventral-to-dorsal (belly-to-back) gradient of BMP activity. The activity of BMP in the ventral region of the embryo suppresses the expression of transcription factors involved in the formation of the central nervous system and segmented muscles. Suppression ensures that these structures are formed only on the dorsal side, where there is decreased activity of BMP.
The final stage of differentiation often involves the formation of several types of differentiated cells from one precursor or stem cellpopulation. Terminal differentiation occurs not only in embryonic development but also in many tissues in postnatal life. Control of this process depends on a system of lateral inhibition in which cells that are differentiating along a particular pathway send out signals that repress similar differentiation by their neighbours. For example, in the developing central nervous system of vertebrates, neurons arise from a simple tube of neuroepithelium, the cells of which possess a surface receptor called Notch. These cells also possess another cell surface molecule called Delta that can bind to and activate Notch on adjacent cells. Activation of Notch initiates a cascade of intracellular events that results in suppression of Delta production and suppression of neuronal differentiation. This means that the neuroepithelium generates only a few cells with high expression of Delta surrounded by a larger number of cells with low expression of Delta. High Delta production and low Notch activation makes the cells develop into neurons. Low Delta production and high Notch activation makes the cells remain as precursor cells or become glial (supporting) cells. A similar mechanism is known to produce the endocrine cells of the pancreas and the goblet cells of the intestinal epithelium. Such lateral inhibition systems work because cells in a population are never quite identical to begin with. There are always small differences, such as in the number of Delta molecules displayed on the cell surface. The mechanism of lateral inhibition amplifies these small differences, using them to bring about differential gene expression that leads to stable and persistent states of cell differentiation.
Three classes of abnormal cell differentiation are dysplasia,metaplasia, and anaplasia. Dysplasia indicates an abnormal arrangement of cells, usually arising from a disturbance in their normal growth behaviour. Some dysplasias are precursor lesions to cancer, whereas others are harmless and regress spontaneously. For example, dysplasia of the uterine cervix, called cervical intraepithelial neoplasia (CIN), may progress to cervical cancer. It can be detected by cervical smear cytology tests (Pap smears).
Metaplasia is the conversion of one cell type into another. In fact, it is not usually the differentiated cells themselves that change but rather the stem cell population from which they are derived. Metaplasia commonly occurs where chronic tissue damage is followed by extensive regeneration. For example, squamous metaplasia of the bronchi occurs when the ciliated respiratory epithelial cells of people who smoke develop into squamous, or flattened, cells. In intestinal metaplasia of the stomach, patches resembling intestinal tissue arise in the gastric mucosa, often in association with gastric ulcers. Both of these types of metaplasia may progress to cancer.
Anaplasia is a loss of visible differentiation that can occur in advanced cancer. In general, early cancers resemble their tissue of origin and are described and classified by their pattern of differentiation. However, as they develop, they produce variants of more abnormal appearance and increased malignancy. Finally, a highly anaplastic growth can occur, in which the cancerous cells bear no visible relation to the parent tissue.
Life on Earth could not exist until a collection of catalysts appeared that could promote the synthesis of more catalysts of the same kind. Early stages in the evolutionary pathway of cells presumably centred on RNA molecules, which not only present specific catalytic surfaces but also contain the potential for their own duplication through the formation of a complementary RNA molecule. It is assumed that a small RNA molecule eventually appeared that was able to catalyze its own duplication.
Imperfections in primitive RNA replication likely gave rise to many variant autocatalytic RNA molecules. Molecules of RNA that acquired variations that increased the speed or the fidelity of self-replication would have outmultiplied other, less-competent RNA molecules. In addition, other small RNA molecules that existed in symbiosis with autocatalytic RNA molecules underwent natural selection for their ability to catalyze useful secondary reactions such as the production of better precursor molecules. In this way, sophisticated families of RNA catalysts could have evolved together, since cooperation between different molecules produced a system that was much more effective at self-replication than a collection of individual RNA catalysts.
Another major step in the evolution of the cell would have been the development, in one family of self-replicating RNA, of a primitive mechanism of protein synthesis. Protein molecules cannot provide the information for the synthesis of other protein molecules like themselves. This information must ultimately be derived from a nucleic acid sequence. Protein synthesis is much more complex than RNA synthesis, and it could not have arisen before a group of powerful RNA catalysts evolved. Each of these catalysts presumably has its counterpart among the RNA molecules that function in the current cell: (1) there was an information RNA molecule, much like messenger RNA (mRNA), whose nucleotide sequence was read to create an amino acid sequence; (2) there was a group of adaptor RNA molecules, much like transfer RNA (tRNA), that could bind to both mRNA and a specific activated amino acid; and (3) finally, there was an RNA catalyst, much like ribosomal RNA (rRNA), that facilitated the joining together of the amino acids aligned on the mRNA by the adaptor RNA.
At some point in the evolution of biological catalysts, the first cell was formed. This would have required the partitioning of the primitive biological catalysts into individual units, each surrounded by a membrane. Membrane formation might have occurred quite simply, since many amphiphilic molecules—half hydrophobic (water-repelling) and half hydrophilic (water-loving)—aggregate to form bilayer sheets in which the hydrophobic portions of the molecules line up in rows to form the interior of the sheet and leave the hydrophilic portions to face the water. Such bilayer sheets can spontaneously close up to form the walls of small, spherical vesicles, as can the phospholipid bilayer membranes of present-day cells.
Lipid structure.
Structure and properties of two representative lipids. Both stearic acid (a fatty acid) and phosphatidylcholine (a phospholipid) are composed of chemical groups that form polar “heads” and nonpolar “tails.” The polar heads are hydrophilic, or soluble in water, whereas the nonpolar tails are hydrophobic, or insoluble in water. Lipid molecules of this composition spontaneously form aggregate structures such as micelles and lipid bilayers, with their hydrophilic ends oriented toward the watery medium and their hydrophobic ends shielded from the water.
As soon as the biological catalysts became compartmentalized into small individual units, or cells, the units would have begun to compete with one another for the same resources. The active competition that ensued must have greatly accelerated evolutionary change, serving as a powerful force for the development of more efficient cells. In this way, cells eventually arose that contained new catalysts, enabling them to use simpler, more abundant precursor molecules for their growth. Because these cells were no longer dependent on preformed ingredients for their survival, they were able to spread far beyond the limited environments where the first primitive cells arose.
It is often assumed that the first cells appeared only after the development of a primitive form of protein synthesis. However, it is by no means certain that cells cannot exist without proteins, and it has been suggested that the first cells contained only RNA catalysts. In either case, protein molecules, with their chemically varied side chains, are more powerful catalysts than RNA molecules; therefore, as time passed, cells arose in which RNA served primarily as genetic material, being directly replicated in each generation and inherited by all progeny cells in order to specify proteins.
As cells became more complex, a need would have arisen for a stabler form of genetic information storage than that provided by RNA. DNA, related to RNA yet chemically stabler, probably appeared rather late in the evolutionary history of cells. Over a period of time, the genetic information in RNA sequences was transferred to DNA sequences, and the ability of RNA molecules to replicate directly was lost. It was only at this point that the central process of biology—the synthesis, one after the other, of DNA, RNA, and protein—appeared.
The first cells presumably resembled prokaryotic cells in lacking nuclei and functional internal compartments, or organelles. These early cells were also anaerobic (not requiring oxygen), deriving their energy from the fermentation of organic molecules that had previously accumulated on the Earth over long periods of time. Eventually, more sophisticated cells evolved that could carry out primitive forms of photosynthesis, in which light energy was harnessed by membrane-bound proteins to form organic molecules with energy-rich chemical bonds. A major turning point in the evolution of life was the development of photosynthesizing prokaryotes requiring only water as an electron donor and capable of producing molecular oxygen. The descendants of these prokaryotes, the blue-green algae (cyanobacteria), still exist as viable life-forms. Their ancestors prospered to such an extent that the atmosphere became rich in the oxygen they produced. The free availability of this oxygen in turn enabled other prokaryotes to evolve aerobic forms of metabolism that were much more efficient in the use of organic molecules as a source of food.
The switch to predominantly aerobic metabolism is thought to have occurred in bacteria approximately 2 billion years ago, about 1.5 billion years after the first cells had formed. Aerobic eukaryotic cells (cells containing nuclei and all the other organelles) probably appeared sometime between 2.1 and 1.5 billion years ago, their lineage having branched off much earlier from that of the prokaryotes. Eukaryotic cells almost certainly became aerobic by engulfing aerobic prokaryotes, with which they lived in a symbiotic relationship. The mitochondria found in both animals and plants are the descendants of such prokaryotes. Later, in branches of the eukaryotic lineage leading to plants and algae, a blue-green algaelike organism was engulfed to perform photosynthesis. It is likely that over a long period of time these organisms became the chloroplasts.
The eukaryotic cell thus apparently arose as an amalgam of different cells, in the process becoming an efficient aerobic cell whose plasma membrane was freed from energy metabolism—one of the major functions of the cell membrane of prokaryotes. The eukaryotic cell membrane was therefore able to become specialized for cell-to-cell communication and cell signaling. It may be partly for this reason that eukaryotic cells were eventually more successful at forming complex multicellular organisms than their simpler prokaryotic relatives.
Antonie van Leeuwenhoek, detail of a portrait by Jan Verkolje; in the Rijksmuseum, Amsterdam.
Robert Brown.
The history of cell theory is a history of the actual observation of cells, because early prediction and speculation about the nature of the cell were generally unsuccessful. The decisive event that allowed the observation of cells was the invention of the microscope in the 16th century, after which interest in the “invisible” world was stimulated. English physicist Robert Hooke, who described cork and other plant tissues in 1665, introduced the term cell because the cellulose walls of dead cork cells reminded him of the blocks of cells occupied by monks. Even after the publication in 1672 of excellent pictures of plant tissues, no significance was attached to the contents within the cell walls. The magnifying powers of the microscope and the inadequacy of techniques for preparing cells for observation precluded a study of the intimate details of the cell contents. The inspired Dutch microscopist Antonie van Leeuwenhoek, beginning in 1673, discovered blood cells,spermatozoa, and a lively world of “animalcules.” A new world of unicellular organisms was opened up. Such discoveries extended the known variety of living things but did not bring insight into their basic uniformity. Moreover, when Leeuwenhoek observed the swarming of his animalcules but failed to observe their division, he could only reinforce the idea that they arose spontaneously.
Cell theory was not formulated for nearly 200 years after the introduction of microscopy. Explanations for this delay range from the poor quality of the microscopes to the persistence of ancient ideas concerning the definition of a fundamental living unit. Many observations of cells were made, but apparently none of the observers was able to assert forcefully that cells are the units of biological structure and function.
Three critical discoveries made during the 1830s, when improved microscopes with suitable lenses, higher powers of magnification without aberration, and more satisfactory illumination became available, were decisive events in the early development of cell theory. First, the nucleus was observed by Scottish botanist Robert Brown in 1833 as a constant component of plant cells. Next, nuclei were also observed and recognized as such in some animal cells. Finally, a living substance called protoplasm was recognized within cells, its vitality made evident by its active streaming, or flowing, movements, especially in plant cells. After these three discoveries, cells, previously considered as mere pores in plant tissue, could no longer be thought of as empty, because they contained living material.
Theodor Schwann.
Matthias Schleiden.
German physiologist Theodor Schwann and German biologist Matthias Schleiden clearly stated in 1839 that cells are the “elementary particles of organisms” in both plants and animals and recognized that some organisms are unicellular and others multicellular. This statement was made in Schwann’s Mikroskopische Untersuchungen über die Übereinstimmung in der Struktur und dem Wachstume der Tiere und Pflanzen (1839; Microscopical Researches into the Accordance in the Structure and Growth of Animals and Plants). Schleiden’s contributions on plants were acknowledged by Schwann as the basis for his comparison of animal and plant structure.
Schleiden and Schwann’s descriptive statements concerning the cellular basis of biologic structure are straightforward and acceptable to modern thought. They recognized the common features of cells to be the membrane, nucleus, and cell body and described them in comparisons of various animal and plant tissues. A statement by Schleiden pointed toward the future direction of cell studies:
“Each cell leads a double life: an independent one, pertaining to its own development alone; and another incidental, insofar as it has become an integral part of a plant. It is, however, easy to perceive that the vital process of the individual cells must form the first, absolutely indispensable fundamental basis, both as regards vegetable physiology and comparative physiology in general.”
Human chromosomes.Schwann and Schleiden were not alone in contributing to this great generalization of natural science, for strong intimations of the cell theory occur in the work of their predecessors. Recognizing that the basic problem was the origin of cells, these early investigators invented a hypothesis of “free cell formation,” according to which cells developed de novo out of an unformed substance, a “cytoblastema,” by a sequence of events in which first the nucleolus develops, followed by the nucleus, the cell body, and finally the cell membrane. The best physical model of the generation of formed bodies then available was crystallization, and their theory was inspired by that model. In retrospect, the hypothesis of free cell formation would not seem to have been justified, however, since cell division, a feature not characteristic of crystallization processes, had frequently been observed by earlier microscopists, especially among single-celled organisms. Even though cell division was observed repeatedly in the following decades, the theory of free cell formation lingered throughout most of the 19th century; however, it came to be thought of more and more as a possible exception to the general principle of the reproduction of cells by division. The correct general principle was affirmed in 1855 by a German pathologist and statesman, Rudolph Virchow, who asserted that “omnis cellula e cellula” (“all cells come from cells”).
Human chromosomes.
The inherently complex events of cell division prevented a quick resolution of the complete sequence of changes that occur during the process. First, it was noted that a cell with a nucleus divides into two cells, each having a nucleus; hence, it was concluded that the nucleus must divide, and direct division of nuclei was duly described by some. Better techniques served to create perplexity, because it was found that during cell division the nucleus as such disappears. Moreover, at the time of division, dimly discerned masses, now recognized as chromosomes, were seen to appear temporarily. Observations in the 1870s culminated in the highly accurate description and interpretation of cell division by German anatomist Walther Flemming in 1882. His advanced techniques of fixing and staining cells enabled him to see that cell reproduction involves the transmission of chromosomes from the parent to daughter cells by the process of mitosis and that the division of the cell body is the terminal event of that reproduction.
The discovery that the number of chromosomes remains constant from one generation to the next resulted in the full description of the process of meiosis. The description of meiosis, combined with the observation that fertilization is fundamentally the union of maternal and paternal sets of chromosomes, culminated in the understanding of the physical basis of reproduction and heredity. Meiosis and fertilization therefore came to be understood as the complementary events in the life cycle of organisms: meiosis halves the number of chromosomes in the formation of spores (plants) or gametes (animals), while fertilization restores the number through the union of gametes. By the 1890s “life” in all of its manifestations could be thought of as an expression of cells.
As the concept of the cell as the elementary particle of life developed during the 19th century, it was paralleled by the “protoplasm” concept—the idea that the protoplasm within the cell is responsible for life. Protoplasm had been defined in 1835 as the ground substance of living material and hence responsible for all living processes. That life is an activity of an elementary particle, the cell, can be contrasted with the view that it is the expression of a living complex substance—even a supermolecule—called a protoplasm. The protoplasm concept was supported by observations of the streaming movements of the apparently slimy contents of living cells.
Advocates of the protoplasm concept implied that cells were either fragments or containers of protoplasm. Suspicious and often contemptuous of information obtained from dead and stained cells, such researchers discovered most of the basic information on the physical properties—mechanical, optical, electrical, and contractile—of the living cell.
An assessment of the usefulness of the concept of protoplasm is difficult. It was not wholly false; on the one hand, it encouraged the study of the chemical and mechanical properties of cell contents, but it also generated a resistance, evident as late as the 1930s, to the development of biochemical techniques for cell fractionation and to the realization that very large molecules (macromolecules) are important cellular constituents. As the cell has become fractionated into its component parts, protoplasm, as a term, no longer has meaning. The word protoplasm is still used, however, in describing the phenomenon of protoplasmic streaming—the phenomenon from which the concept of protoplasm originally emerged.
French chemist and microbiologist Louis Pasteur performing a scientific experiment. Pasteur's studies helped establish the principle of biogenesis, the development of new organisms from the reproduction of other organisms.
Appreciation of the cell as the unit of life has accrued from important sources other than microscopy; perhaps the most important is microbiology. Even though the small size of microorganisms prohibited much observation of their detailed structure until the advent of electron microscopy, they could be grown easily and rapidly. Thus it was that French chemist and microbiologist Louis Pasteur’s studies of microbes published in 1861 helped to establish the principle of biogenesis—namely, that organisms arise only by the reproduction of other organisms. Fundamental ideas regarding the metabolic attributes of cells—that is, their ability to transform simple nutritional substances into cell substance and utilizable energy—came from microbiology. Pasteur perhaps overplayed the relation between catalysis and the living state of cells in considering enzymatic action to be an attribute of the living cell rather than of the catalytic molecules (enzymes) contained in the cell; it is a fact, however, that much of cell chemistry is enzyme chemistry—and that enzymes are one defining attribute of cells. The techniques of microbiology eventually opened the way for microbial genetics, which in turn provided the means for solving the fundamental problems of molecular biology that were inaccessible at first to direct attack by biochemical methods.
The science of molecular biology would be most capable of overthrowing the cell theory if the latter were an exaggerated generalization. On the contrary, molecular biology has become the foundation of cell science, for it has demonstrated not only that basic processes such as the genetic code and protein synthesis are similar in all living systems but also that they are made possible by the same cell components—e.g., chromosomes, ribosomes, and membranes.
In the overlapping histories of cell biology and medicine, two events are especially important. One, the identification in 1827 by Prussian-Estonian embryologist Karl Ernst Ritter von Baer of the ovum (unfertilized egg) as a cell, was important considering the many ways it often differs from other cells. Baer not only laid the foundations for reproductive biology but also provided important evidence for the cell theory at a critical time. The second important event was the promotion in 1855 of the concept of “cellular pathology” by Virchow. His idea that human diseases are diseases of cells and can be identified and understood as such gave an authority to cell theory.
Karl Ernst, Ritter von Baer. Karl Ernst, Ritter von Baer, detail of a lithograph by Rudolf Hoffmann, 1839.
Although biochemistry might have made considerable progress without cell theory, each influenced the other almost from the start. When it was established that most biochemical phenomena are shared by all cells, the cell could be defined by its metabolism as well as by its structure. Cytochemistry, or histochemistry, made a brilliant start in 1869, when Swiss biochemist Johann Friedrich Miescher postulated that the nucleus must have a characteristic chemistry and then went on to discover nucleic acids, which have since been shown to be the crucial molecules of inheritance and metabolism.
Cell theory by itself cannot explain the development and unity of the multicellular organism. A cell is not necessarily an independently functioning unit, and a plant or an animal is not merely an accumulation of individual cells. Fortunately, however, the long controversy centring upon the individuality and separateness of cells has ended. Cell biology now focuses on the interactions and communication among cells as well as on the analysis of the single cell. The influence of the environment on the cell has always been considered important; now it has been recognized that one important part of the environment of a cell is other cells.
Cell theory thus is not so comprehensive as to eliminate the concept of the organism as more than the sum of its parts. But the study of a particular organism requires the investigation of cells both as individuals and as groups. The problem of cancer is an example: a plant or animal governs the division of its own cells; the right cells must divide, be differentiated, and then be integrated into the proper organ system at the right time and place. Breakdown results in a variety of abnormalities, one of which is cancer. When the cell biologist studies the problem of the regulation of cell division, the ultimate objective is to understand the effect of the whole organism on an individual cell.
The cell (from Latincella, meaning "small room") is the basic structural, functional, and biological unit of all known organisms. A cell is the smallest unit of life. Cells are often called the "building blocks of life". The study of cells is called cell biology, cellular biology, or cytology.
The number of cells in plants and animals varies from species to species; it has been estimated that humans contain somewhere around 40 trillion (4×1013) cells. The human brain accounts for around 80 billion of these cells.
Cells were discovered by Robert Hooke in 1665, {?} who named them for their resemblance to cells inhabited by Christian monks in a monastery. Cell theory, first developed in 1839 by Matthias Jakob Schleiden and Theodor Schwann, states that all organisms are composed of one or more cells, that cells are the fundamental unit of structure and function in all living organisms, and that all cells come from pre-existing cells. Cells emerged on Earth at least 3.5 billion years ago.
The cells of eukaryotes (left) and prokaryotes (right).
Enclosing the cell is the cell envelope – generally consisting of a plasma membrane covered by a cell wall which, for some bacteria, may be further covered by a third layer called a capsule. Though most prokaryotes have both a cell membrane and a cell wall, there are exceptions such as Mycoplasma (bacteria) and Thermoplasma (archaea) which only possess the cell membrane layer. The envelope gives rigidity to the cell and separates the interior of the cell from its environment, serving as a protective filter. The cell wall consists of peptidoglycan in bacteria, and acts as an additional barrier against exterior forces. It also prevents the cell from expanding and bursting (cytolysis) from osmotic pressure due to a hypotonic environment. Some eukaryotic cells (plant cells and fungal cells) also have a cell wall.
Inside the cell is the cytoplasmic region that contains the genome (DNA), ribosomes and various sorts of inclusions. The genetic material is freely found in the cytoplasm. Prokaryotes can carry extrachromosomal DNA elements called plasmids, which are usually circular. Linear bacterial plasmids have been identified in several species of spirochete bacteria, including members of the genus Borrelia notably Borrelia burgdorferi, which causes Lyme disease. Though not forming a nucleus, the DNA is condensed in a nucleoid. Plasmids encode additional genes, such as antibiotic resistance genes.
On the outside, flagella and pili project from the cell's surface. These are structures (not present in all prokaryotes) made of proteins that facilitate movement and communication between cells.
Plants, animals,fungi,slime moulds,protozoa, and algae are all eukaryotic. These cells are about fifteen times wider than a typical prokaryote and can be as much as a thousand times greater in volume. The main distinguishing feature of eukaryotes as compared to prokaryotes is compartmentalization: the presence of membrane-bound organelles (compartments) in which specific activities take place. Most important among these is a cell nucleus, an organelle that houses the cell's DNA. This nucleus gives the eukaryote its name, which means "true kernel (nucleus)". Other differences include:
The plasma membrane resembles that of prokaryotes in function, with minor differences in the setup. Cell walls may or may not be present.
The eukaryotic DNA is organized in one or more linear molecules, called chromosomes, which are associated with histone proteins. All chromosomal DNA is stored in the cell nucleus, separated from the cytoplasm by a membrane. Some eukaryotic organelles such as mitochondria also contain some DNA.
Many eukaryotic cells are ciliated with primary cilia. Primary cilia play important roles in chemosensation, mechanosensation, and thermosensation. Each cilium may thus be "viewed as a sensory cellular antennae that coordinates a large number of cellular signaling pathways, sometimes coupling the signaling to ciliary motility or alternatively to cell division and differentiation."
All cells, whether prokaryotic or eukaryotic, have a membrane that envelops the cell, regulates what moves in and out (selectively permeable), and maintains the electric potential of the cell. Inside the membrane, the cytoplasm takes up most of the cell's volume. All cells (except red blood cells which lack a cell nucleus and most organelles to accommodate maximum space for hemoglobin) possess DNA, the hereditary material of genes, and RNA, containing the information necessary to build various proteins such as enzymes, the cell's primary machinery. There are also other kinds of biomolecules in cells. This article lists these primary cellular components, then briefly describes their function.
The cell membrane, or plasma membrane, is a biological membrane that surrounds the cytoplasm of a cell. In animals, the plasma membrane is the outer boundary of the cell, while in plants and prokaryotes it is usually covered by a cell wall. This membrane serves to separate and protect a cell from its surrounding environment and is made mostly from a double layer of phospholipids, which are amphiphilic (partly hydrophobic and partly hydrophilic). Hence, the layer is called a phospholipid bilayer, or sometimes a fluid mosaic membrane. Embedded within this membrane is a macromolecular structure called the porosome the universal secretory portal in cells and a variety of protein molecules that act as channels and pumps that move different molecules into and out of the cell. The membrane is semi-permeable, and selectively permeable, in that it can either let a substance (molecule or ion) pass through freely, pass through to a limited extent or not pass through at all. Cell surface membranes also contain receptor proteins that allow cells to detect external signaling molecules such as hormones.
The cytoskeleton acts to organize and maintain the cell's shape; anchors organelles in place; helps during endocytosis, the uptake of external materials by a cell, and cytokinesis, the separation of daughter cells after cell division; and moves parts of the cell in processes of growth and mobility. The eukaryotic cytoskeleton is composed of microfilaments,intermediate filaments and microtubules. There are a great number of proteins associated with them, each controlling a cell's structure by directing, bundling, and aligning filaments. The prokaryotic cytoskeleton is less well-studied but is involved in the maintenance of cell shape, polarity and cytokinesis. The subunit protein of microfilaments is a small, monomeric protein called actin. The subunit of microtubules is a dimeric molecule called tubulin. Intermediate filaments are heteropolymers whose subunits vary among the cell types in different tissues. But some of the subunit protein of intermediate filaments include vimentin,desmin,lamin (lamins A, B and C), keratin (multiple acidic and basic keratins), neurofilament proteins (NF–L, NF–M).
A fluorescent image of an endothelial cell. Nuclei are stained blue, mitochondria are stained red, and microfilaments are stained green.
Two different kinds of genetic material exist: deoxyribonucleic acid (DNA) and ribonucleic acid (RNA). Cells use DNA for their long-term information storage. The biological information contained in an organism is encoded in its DNA sequence. RNA is used for information transport (e.g., mRNA) and enzymatic functions (e.g., ribosomal RNA). Transfer RNA (tRNA) molecules are used to add amino acids during protein translation.
A human cell has genetic material contained in the cell nucleus (the nuclear genome) and in the mitochondria (the mitochondrial genome). In humans the nuclear genome is divided into 46 linear DNA molecules called chromosomes, including 22 homologous chromosome pairs and a pair of sex chromosomes. The mitochondrial genome is a circular DNA molecule distinct from the nuclear DNA. Although the mitochondrial DNA is very small compared to nuclear chromosomes, it codes for 13 proteins involved in mitochondrial energy production and specific tRNAs.
Foreign genetic material (most commonly DNA) can also be artificially introduced into the cell by a process called transfection. This can be transient, if the DNA is not inserted into the cell's genome, or stable, if it is. Certain viruses also insert their genetic material into the genome.
Organelles are parts of the cell which are adapted and/or specialized for carrying out one or more vital functions, analogous to the organs of the human body (such as the heart, lung, and kidney, with each organ performing a different function). Both eukaryotic and prokaryotic cells have organelles, but prokaryotic organelles are generally simpler and are not membrane-bound.
There are several types of organelles in a cell. Some (such as the nucleus and golgi apparatus) are typically solitary, while others (such as mitochondria,chloroplasts,peroxisomes and lysosomes) can be numerous (hundreds to thousands). The cytosol is the gelatinous fluid that fills the cell and surrounds the organelles.
Eukaryotic
Human cancer cells, specifically HeLa cells, with DNA stained blue. The central and rightmost cell are in interphase, so their DNA is diffuse and the entire nuclei are labelled. The cell on the left is going through mitosis and its chromosomes have condensed.
Cell nucleus: A cell's information center, the cell nucleus is the most conspicuous organelle found in a eukaryotic cell. It houses the cell's chromosomes, and is the place where almost all DNA replication and RNA synthesis (transcription) occur. The nucleus is spherical and separated from the cytoplasm by a double membrane called the nuclear envelope. The nuclear envelope isolates and protects a cell's DNA from various molecules that could accidentally damage its structure or interfere with its processing. During processing, DNA is transcribed, or copied into a special RNA, called messenger RNA (mRNA). This mRNA is then transported out of the nucleus, where it is translated into a specific protein molecule. The nucleolus is a specialized region within the nucleus where ribosome subunits are assembled. In prokaryotes, DNA processing takes place in the cytoplasm.
Mitochondria and Chloroplasts: generate energy for the cell. Mitochondria are self-replicating organelles that occur in various numbers, shapes, and sizes in the cytoplasm of all eukaryotic cells. Respiration occurs in the cell mitochondria, which generate the cell's energy by oxidative phosphorylation, using oxygen to release energy stored in cellular nutrients (typically pertaining to glucose) to generate ATP. Mitochondria multiply by binary fission, like prokaryotes. Chloroplasts can only be found in plants and algae, and they capture the sun's energy to make carbohydrates through photosynthesis.
Endoplasmic reticulum: The endoplasmic reticulum (ER) is a transport network for molecules targeted for certain modifications and specific destinations, as compared to molecules that float freely in the cytoplasm. The ER has two forms: the rough ER, which has ribosomes on its surface that secrete proteins into the ER, and the smooth ER, which lacks ribosomes. The smooth ER plays a role in calcium sequestration and release.
Golgi apparatus: The primary function of the Golgi apparatus is to process and package the macromolecules such as proteins and lipids that are synthesized by the cell.
Centrosome: the cytoskeleton organiser: The centrosome produces the microtubules of a cell – a key component of the cytoskeleton. It directs the transport through the ER and the Golgi apparatus. Centrosomes are composed of two centrioles, which separate during cell division and help in the formation of the mitotic spindle. A single centrosome is present in the animal cells. They are also found in some fungi and algae cells.
Vacuoles: Vacuoles sequester waste products and in plant cells store water. They are often described as liquid filled space and are surrounded by a membrane. Some cells, most notably Amoeba, have contractile vacuoles, which can pump water out of the cell if there is too much water. The vacuoles of plant cells and fungal cells are usually larger than those of animal cells.
Eukaryotic and prokaryotic
Ribosomes: The ribosome is a large complex of RNA and protein molecules. They each consist of two subunits, and act as an assembly line where RNA from the nucleus is used to synthesise proteins from amino acids. Ribosomes can be found either floating freely or bound to a membrane (the rough endoplasmatic reticulum in eukaryotes, or the cell membrane in prokaryotes).
Many cells also have structures which exist wholly or partially outside the cell membrane. These structures are notable because they are not protected from the external environment by the semipermeable cell membrane. In order to assemble these structures, their components must be carried across the cell membrane by export processes.
Many types of prokaryotic and eukaryotic cells have a cell wall. The cell wall acts to protect the cell mechanically and chemically from its environment, and is an additional layer of protection to the cell membrane. Different types of cell have cell walls made up of different materials; plant cell walls are primarily made up of cellulose, fungi cell walls are made up of chitin and bacteria cell walls are made up of peptidoglycan.
Flagella are organelles for cellular mobility. The bacterial flagellum stretches from cytoplasm through the cell membrane(s) and extrudes through the cell wall. They are long and thick thread-like appendages, protein in nature. A different type of flagellum is found in archaea and a different type is found in eukaryotes.
Fimbriae
A fimbria (plural fimbriae also known as a pilus, plural pili) is a short, thin, hair-like filament found on the surface of bacteria. Fimbriae are formed of a protein called pilin (antigenic) and are responsible for the attachment of bacteria to specific receptors on human cells (cell adhesion). There are special types of pili involved in bacterial conjugation.
DNA replication, or the process of duplicating a cell's genome, always happens when a cell divides through mitosis or binary fission. This occurs during the S phase of the cell cycle.
In meiosis, the DNA is replicated only once, while the cell divides twice. DNA replication only occurs before meiosis I. DNA replication does not occur when the cells divide the second time, in meiosis II. Replication, like all cellular activities, requires specialized proteins for carrying out the job.
In general, cells of all organisms contain enzyme systems that scan their DNA for damages and carry out repair processes when damages are detected. Diverse repair processes have evolved in organisms ranging from bacteria to humans. The widespread prevalence of these repair processes indicates the importance of maintaining cellular DNA in an undamaged state in order to avoid cell death or errors of replication due to damages that could lead to mutation.E. coli bacteria are a well-studied example of a cellular organism with diverse well-defined DNA repair processes. These include: (1) nucleotide excision repair, (2) DNA mismatch repair, (3) non-homologous end joining of double-strand breaks, (4) recombinational repair and (5) light-dependent repair (photoreactivation).
An overview of protein synthesis.
Within the nucleus of the cell (light blue), genes (DNA, dark blue) are transcribed into RNA. This RNA is then subject to post-transcriptional modification and control, resulting in a mature mRNA (red) that is then transported out of the nucleus and into the cytoplasm (peach), where it undergoes translation into a protein. mRNA is translated by ribosomes (purple) that match the three-base codons of the mRNA to the three-base anti-codons of the appropriate tRNA. Newly synthesized proteins (black) are often further modified, such as by binding to an effector molecule (orange), to become fully active..
Between successive cell divisions, cells grow through the functioning of cellular metabolism. Cell metabolism is the process by which individual cells process nutrient molecules. Metabolism has two distinct divisions: catabolism, in which the cell breaks down complex molecules to produce energy and reducing power, and anabolism, in which the cell uses energy and reducing power to construct complex molecules and perform other biological functions. Complex sugars consumed by the organism can be broken down into simpler sugar molecules called monosaccharides such as glucose. Once inside the cell, glucose is broken down to make adenosine triphosphate (ATP), a molecule that possesses readily available energy, through two different pathways.
Cells are capable of synthesizing new proteins, which are essential for the modulation and maintenance of cellular activities. This process involves the formation of new protein molecules from amino acid building blocks based on information encoded in DNA/RNA. Protein synthesis generally consists of two major steps: transcription and translation.
Transcription is the process where genetic information in DNA is used to produce a complementary RNA strand. This RNA strand is then processed to give messenger RNA (mRNA), which is free to migrate through the cell. mRNA molecules bind to protein-RNA complexes called ribosomes located in the cytosol, where they are translated into polypeptide sequences. The ribosome mediates the formation of a polypeptide sequence based on the mRNA sequence. The mRNA sequence directly relates to the polypeptide sequence by binding to transfer RNA (tRNA) adapter molecules in binding pockets within the ribosome. The new polypeptide then folds into a functional three-dimensional protein molecule.
Unicellular organisms can move in order to find food or escape predators. Common mechanisms of motion include flagella and cilia.
In multicellular organisms, cells can move during processes such as wound healing, the immune response and cancer metastasis. For example, in wound healing in animals, white blood cells move to the wound site to kill the microorganisms that cause infection. Cell motility involves many receptors, crosslinking, bundling, binding, adhesion, motor and other proteins. The process is divided into three steps – protrusion of the leading edge of the cell, adhesion of the leading edge and de-adhesion at the cell body and rear, and cytoskeletal contraction to pull the cell forward. Each step is driven by physical forces generated by unique segments of the cytoskeleton.
In complex multicellular organisms, cells specialize into different cell types that are adapted to particular functions. In mammals, major cell types include skin cells,muscle cells,neurons,blood cells,fibroblasts,stem cells, and others. Cell types differ both in appearance and function, yet are genetically identical. Cells are able to be of the same genotype but of different cell type due to the differential expression of the genes they contain.
Most distinct cell types arise from a single totipotent cell, called a zygote, that differentiates into hundreds of different cell types during the course of development. Differentiation of cells is driven by different environmental cues (such as cell–cell interaction) and intrinsic differences (such as those caused by the uneven distribution of molecules during division).
The first evidence of multicellularity is from cyanobacteria-like organisms that lived between 3 and 3.5 billion years ago. Other early fossils of multicellular organisms include the contested Grypania spiralis and the fossils of the black shales of the PalaeoproterozoicFrancevillian Group Fossil B Formation in Gabon.
The evolution of multicellularity from unicellular ancestors has been replicated in the laboratory, in evolution experiments using predation as the selective pressure.
Stromatolites are left behind by cyanobacteria, also called blue-green algae. They are the oldest known fossils of life on Earth. This one-billion-year-old fossil is from Glacier National Park in the United States.
There are several theories about the origin of small molecules that led to life on the early Earth. They may have been carried to Earth on meteorites (see Murchison meteorite), created at deep-sea vents, or synthesized by lightning in a reducing atmosphere (see Miller–Urey experiment). There is little experimental data defining what the first self-replicating forms were. RNA is thought to be the earliest self-replicating molecule, as it is capable of both storing genetic information and catalyzing chemical reactions (see RNA world hypothesis), but some other entity with the potential to self-replicate could have preceded RNA, such as clay or peptide nucleic acid.
Cells emerged at least 3.5 billion years ago. The current belief is that these cells were heterotrophs. The early cell membranes were probably more simple and permeable than modern ones, with only a single fatty acid chain per lipid. Lipids are known to spontaneously form bilayered vesicles in water, and could have preceded RNA, but the first cell membranes could also have been produced by catalytic RNA, or even have required structural proteins before they could form.
There is still considerable debate about whether organelles like the hydrogenosome predated the origin of mitochondria, or vice versa: see the hydrogen hypothesis for the origin of eukaryotic cells.
1665: Robert Hooke discovered cells in cork, then in living plant tissue using an early compound microscope. He coined the term cell (from Latincella, meaning "small room") in his book Micrographia (1665).
1839: Theodor Schwann and Matthias Jakob Schleiden elucidated the principle that plants and animals are made of cells, concluding that cells are a common unit of structure and development, and thus founding the cell theory.
1855: Rudolf Virchow stated that new cells come from pre-existing cells by cell division (omnis cellula ex cellula).
1859: The belief that life forms can occur spontaneously (generatio spontanea) was contradicted by Louis Pasteur (1822–1895) (although Francesco Redi had performed an experiment in 1668 that suggested the same conclusion).


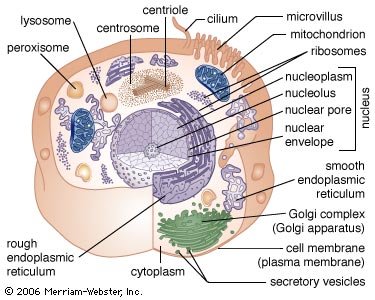
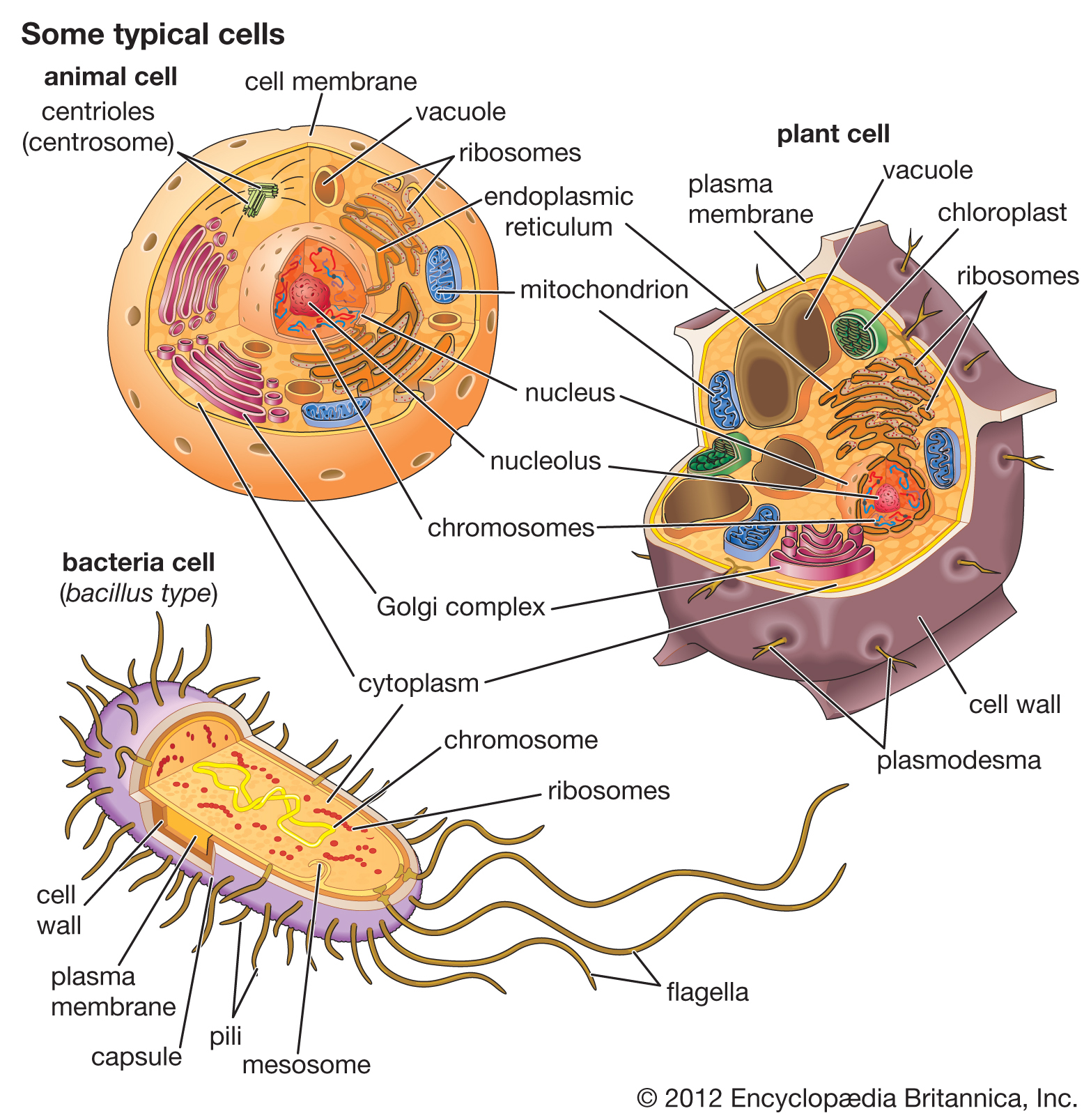
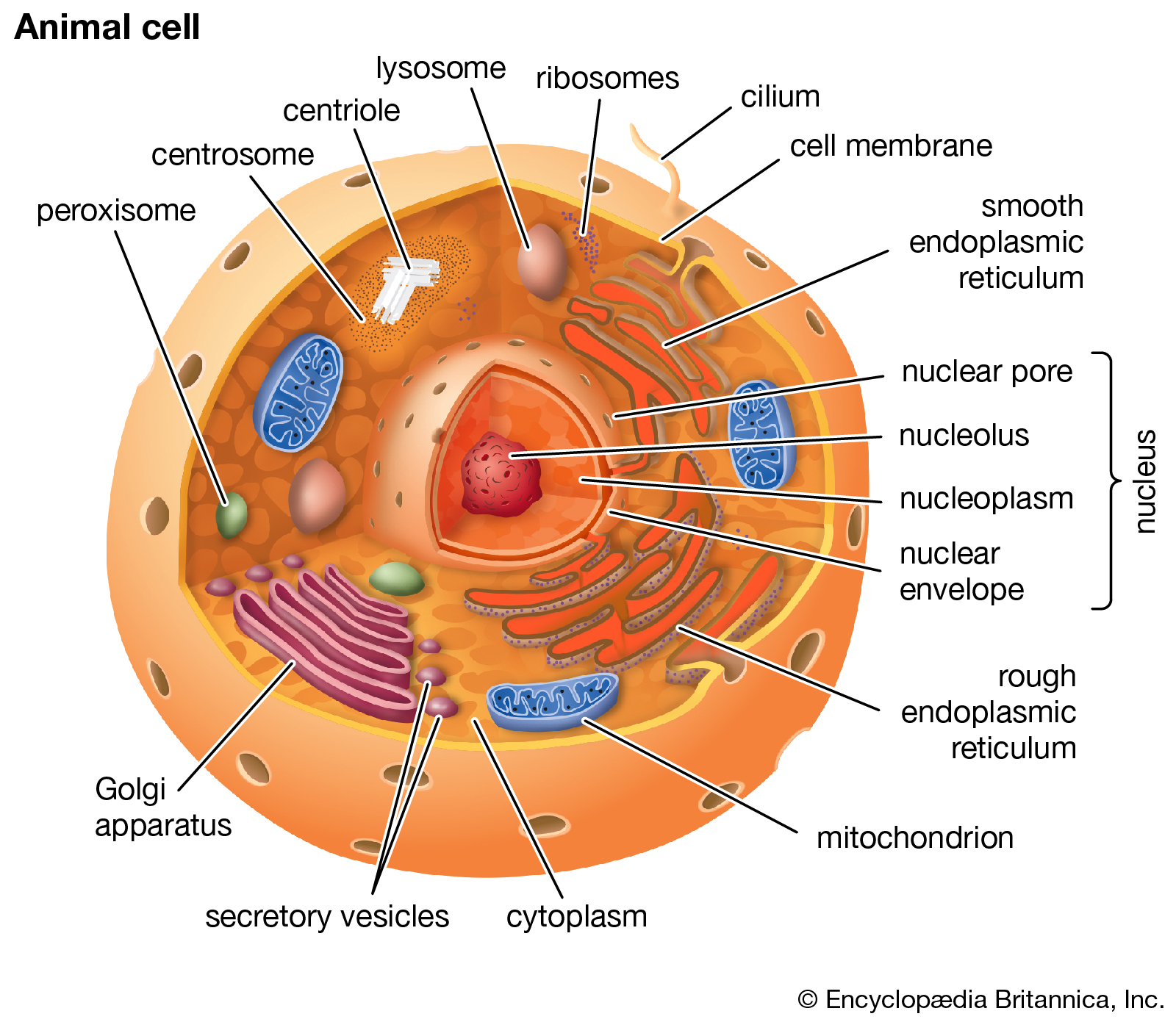
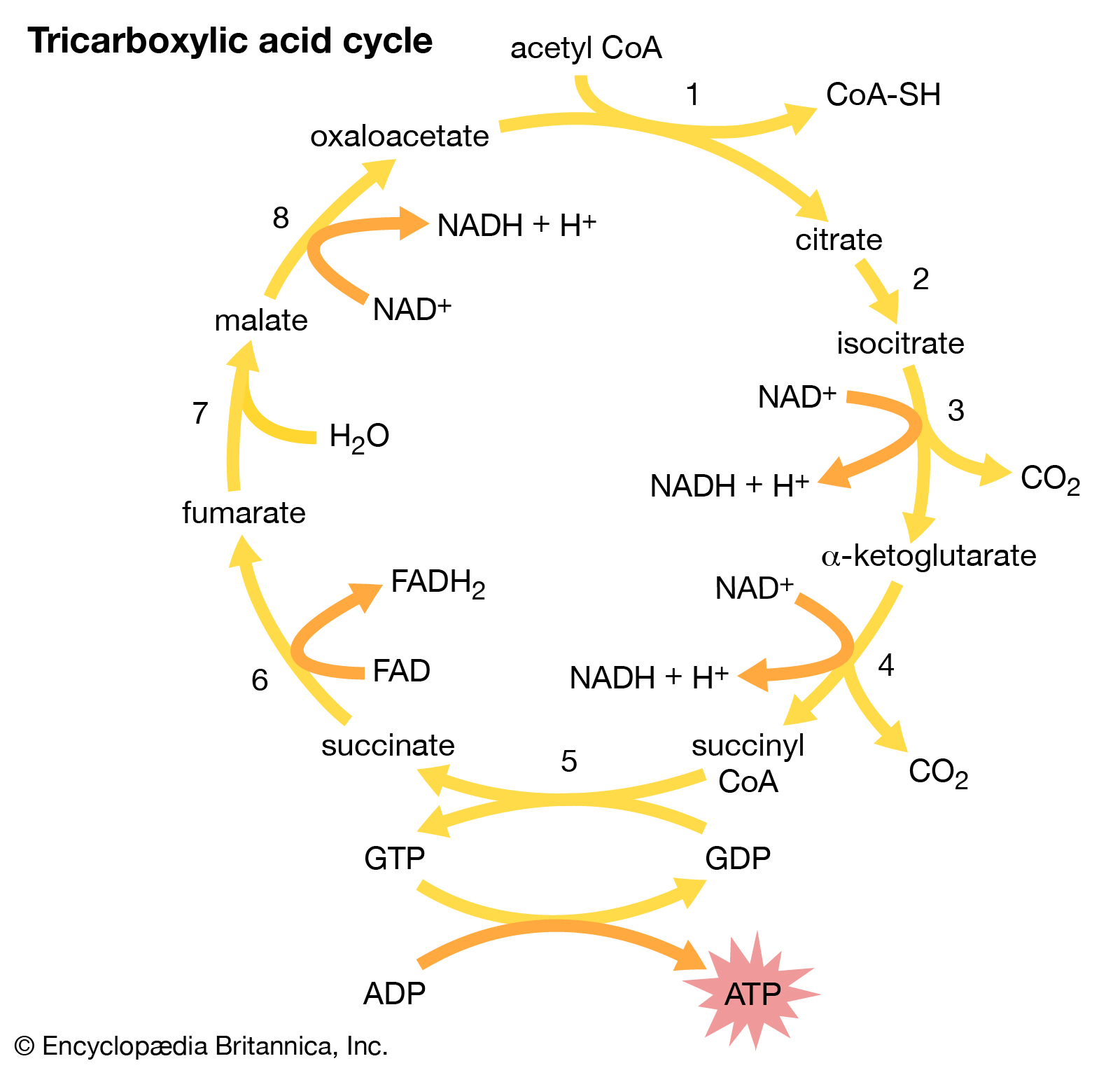
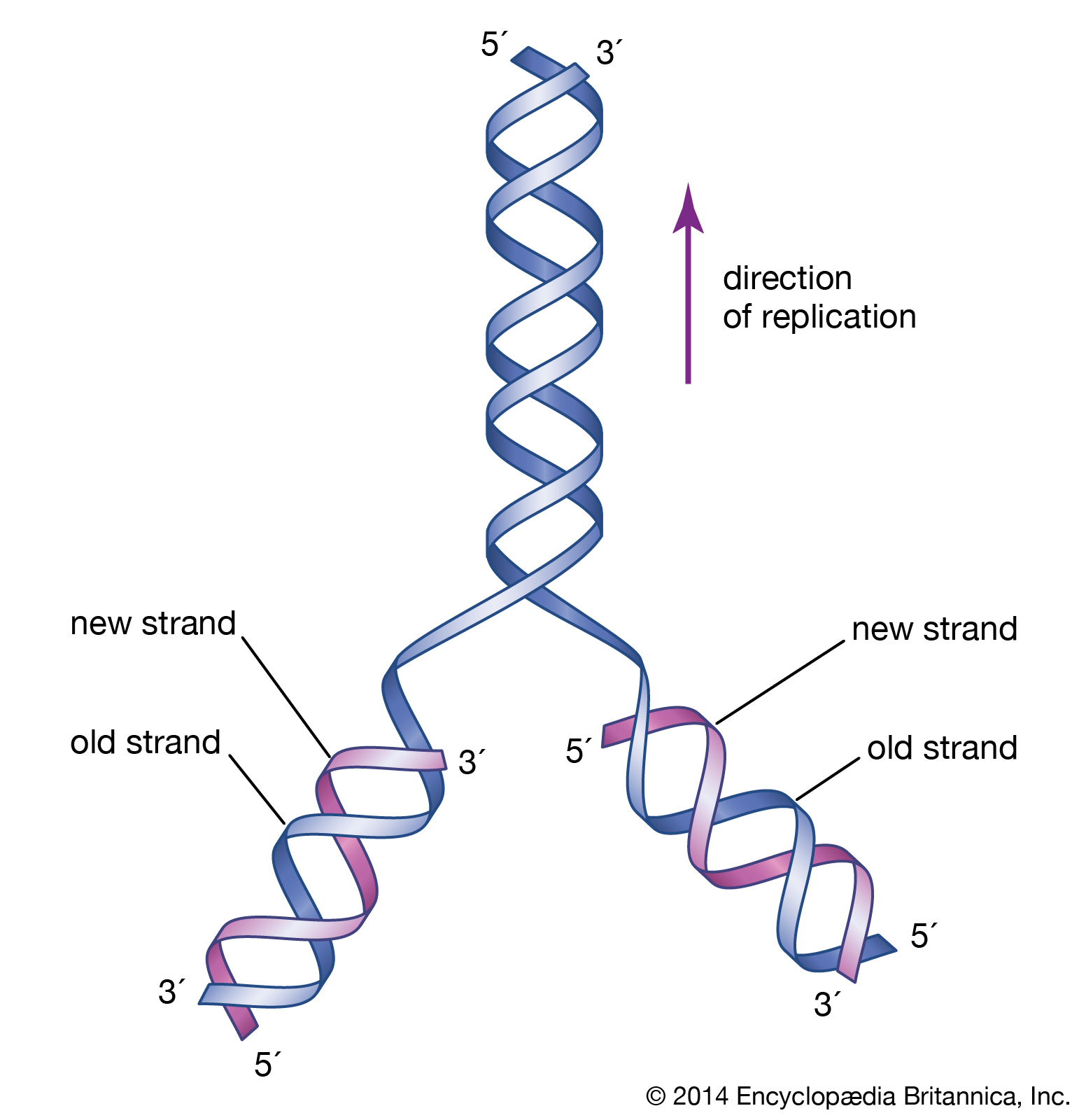
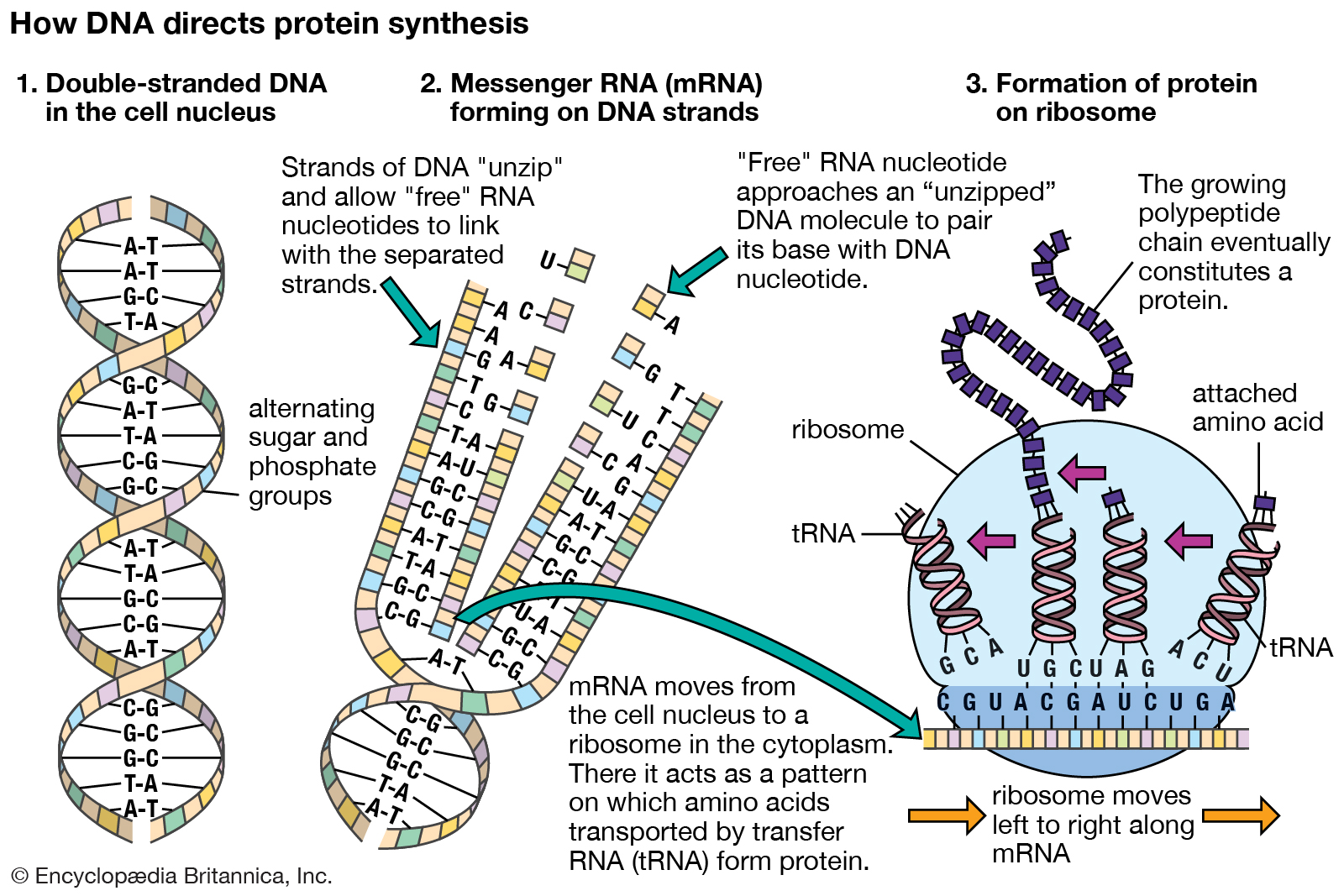
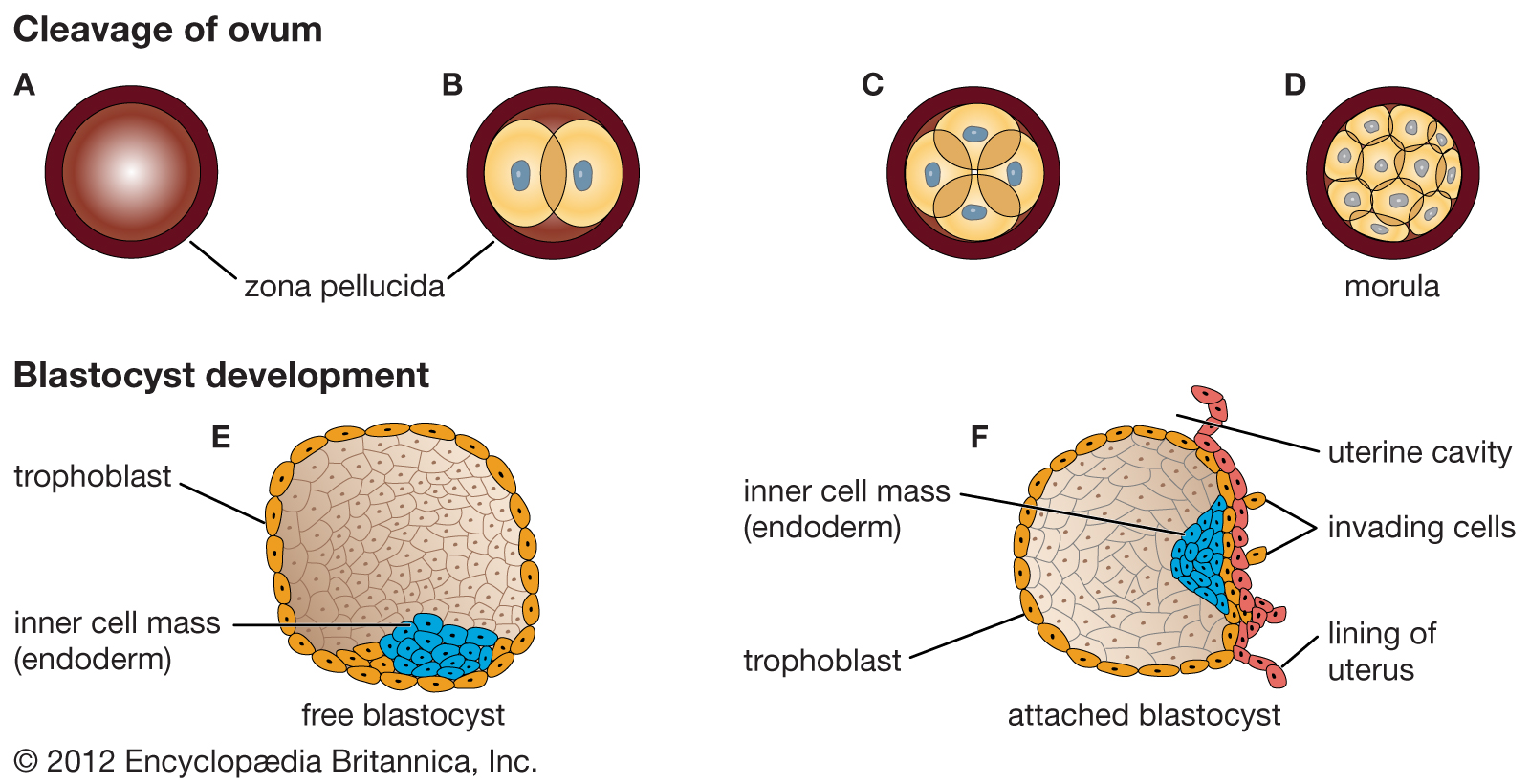
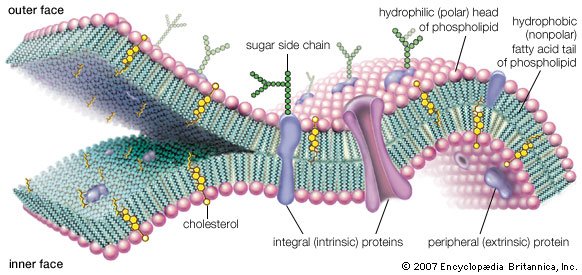
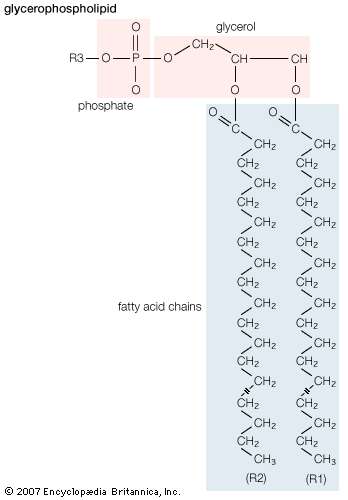
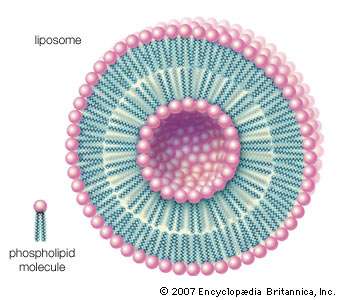

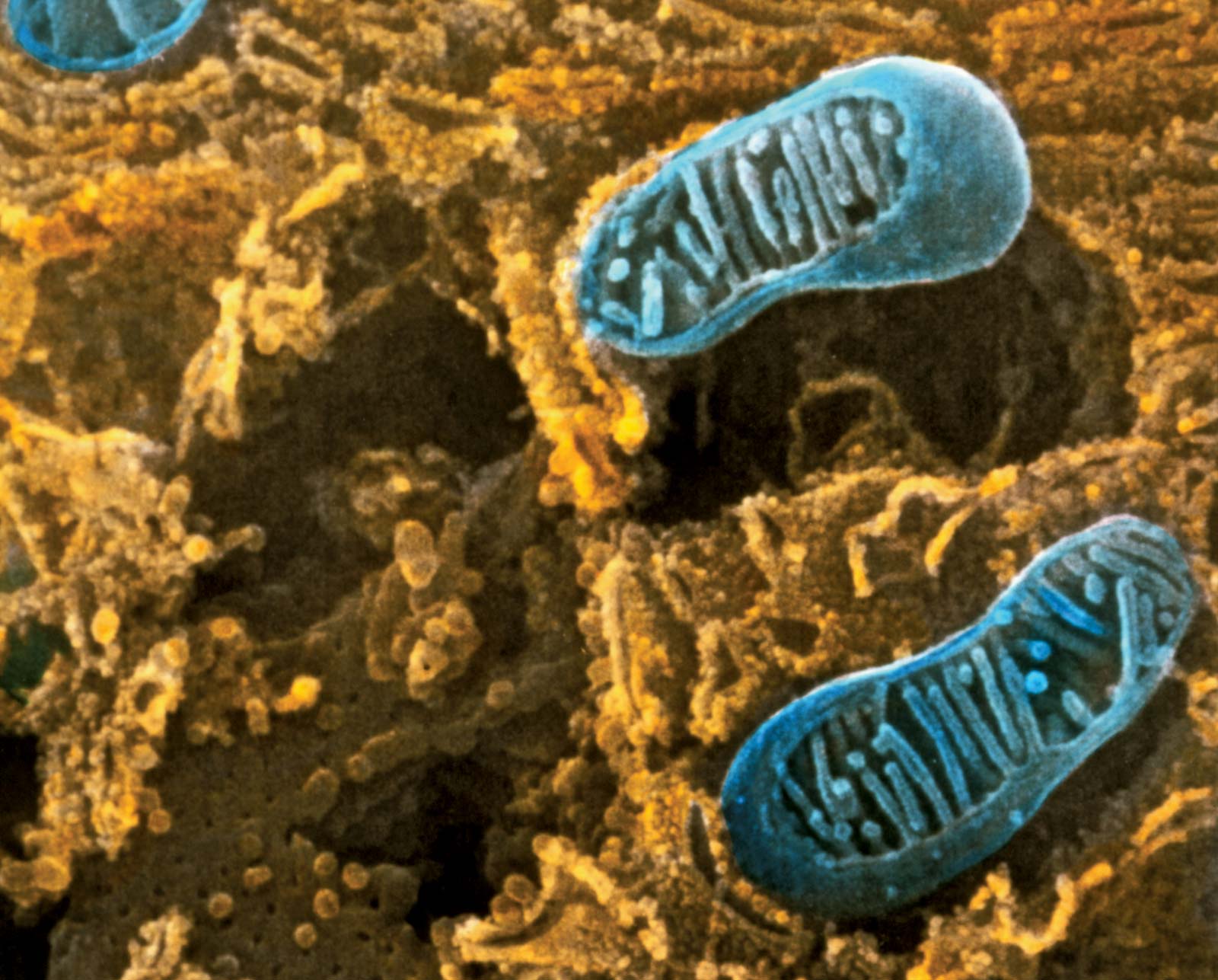
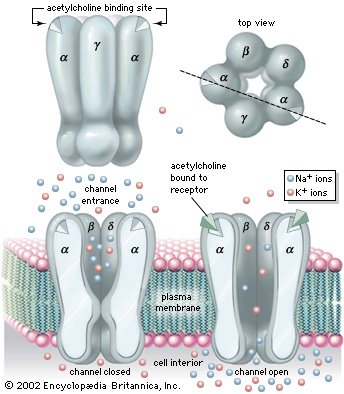
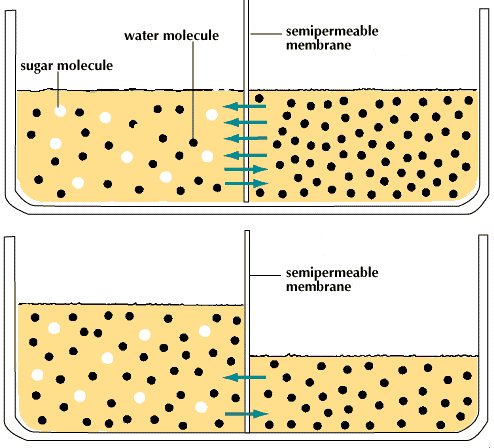
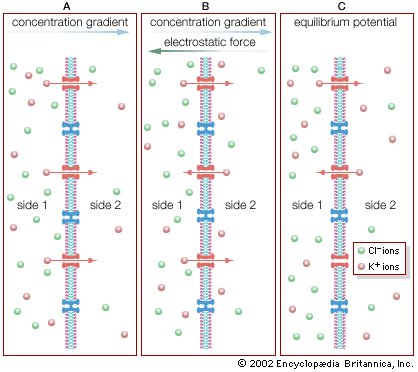
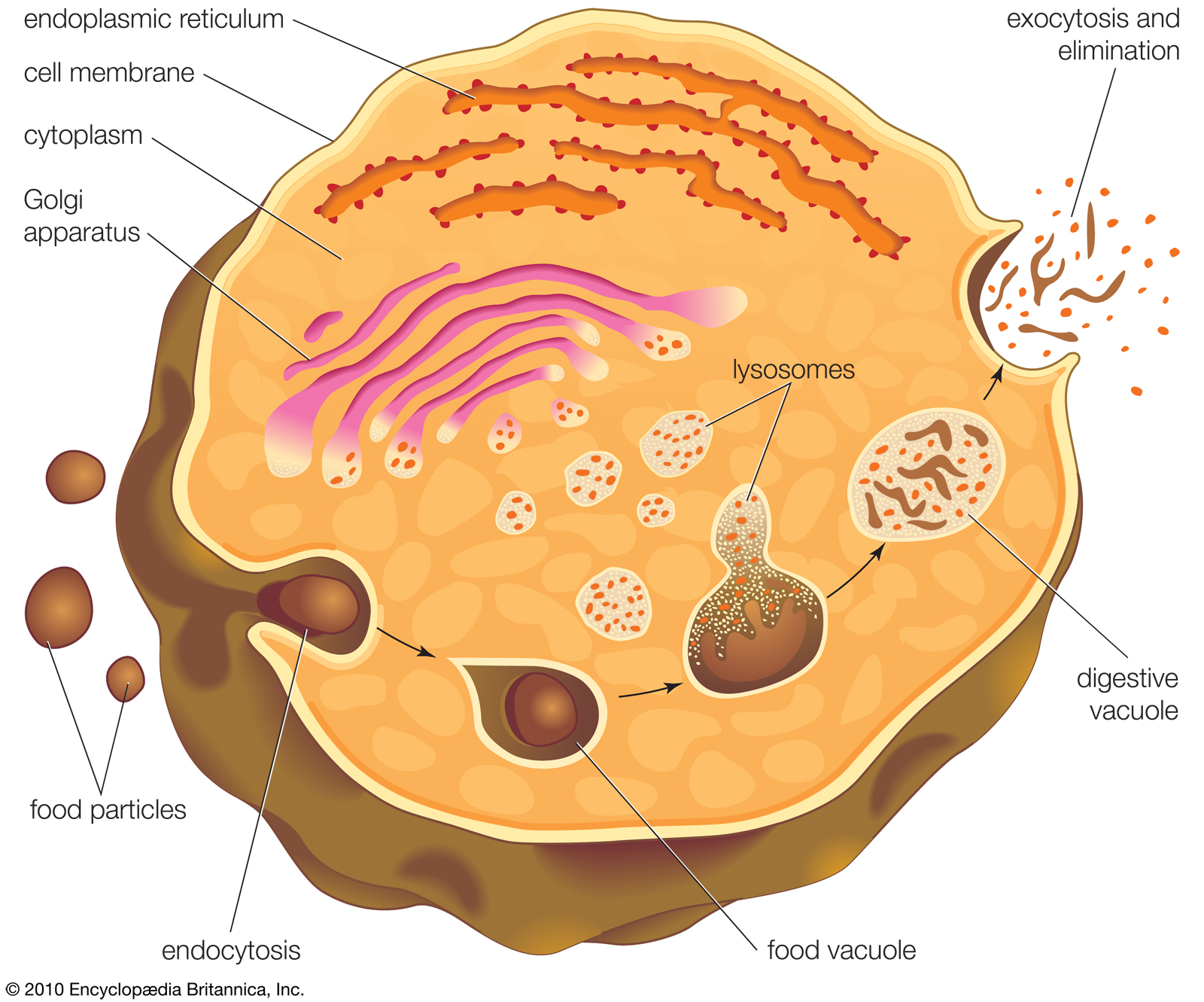
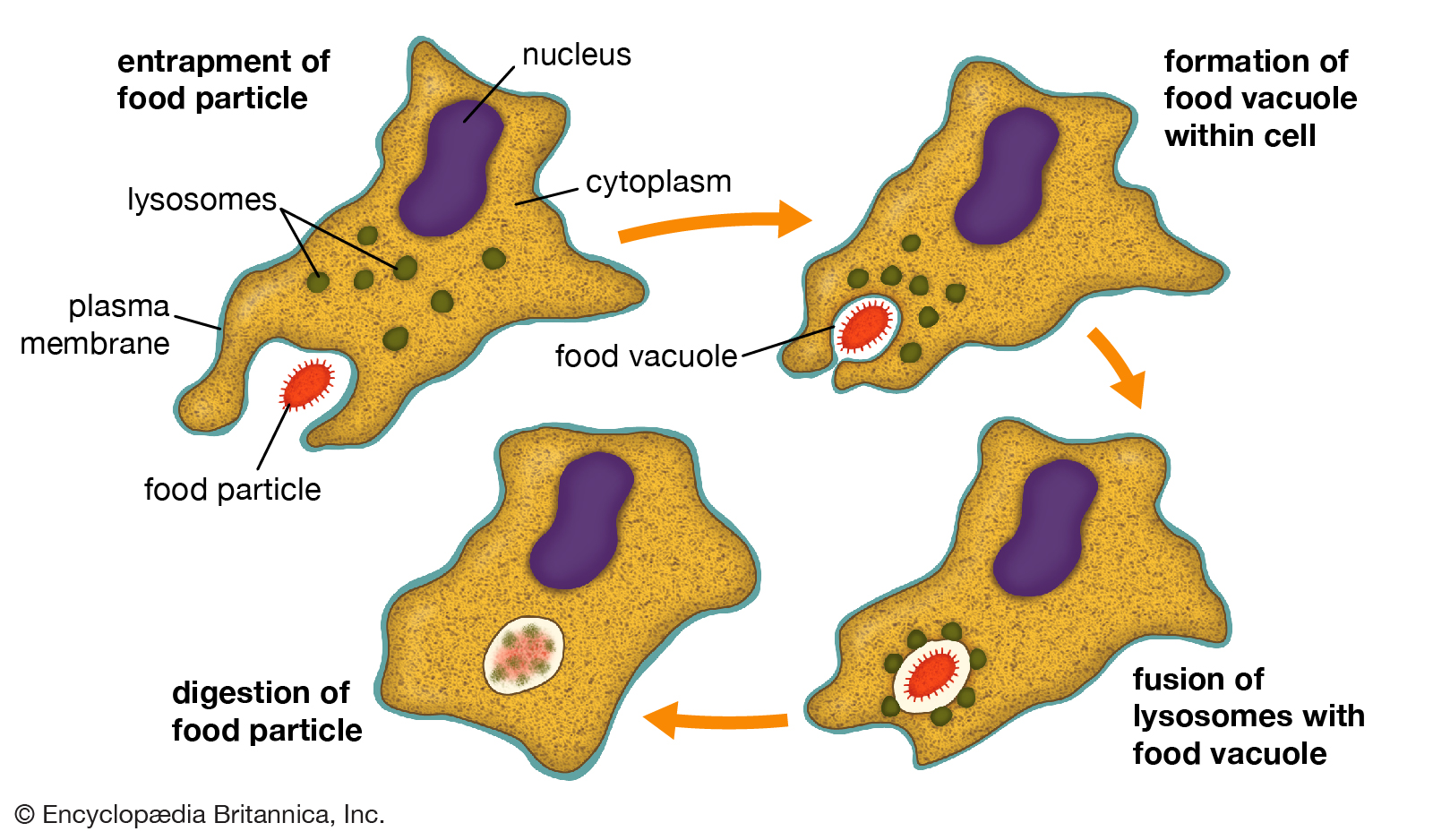
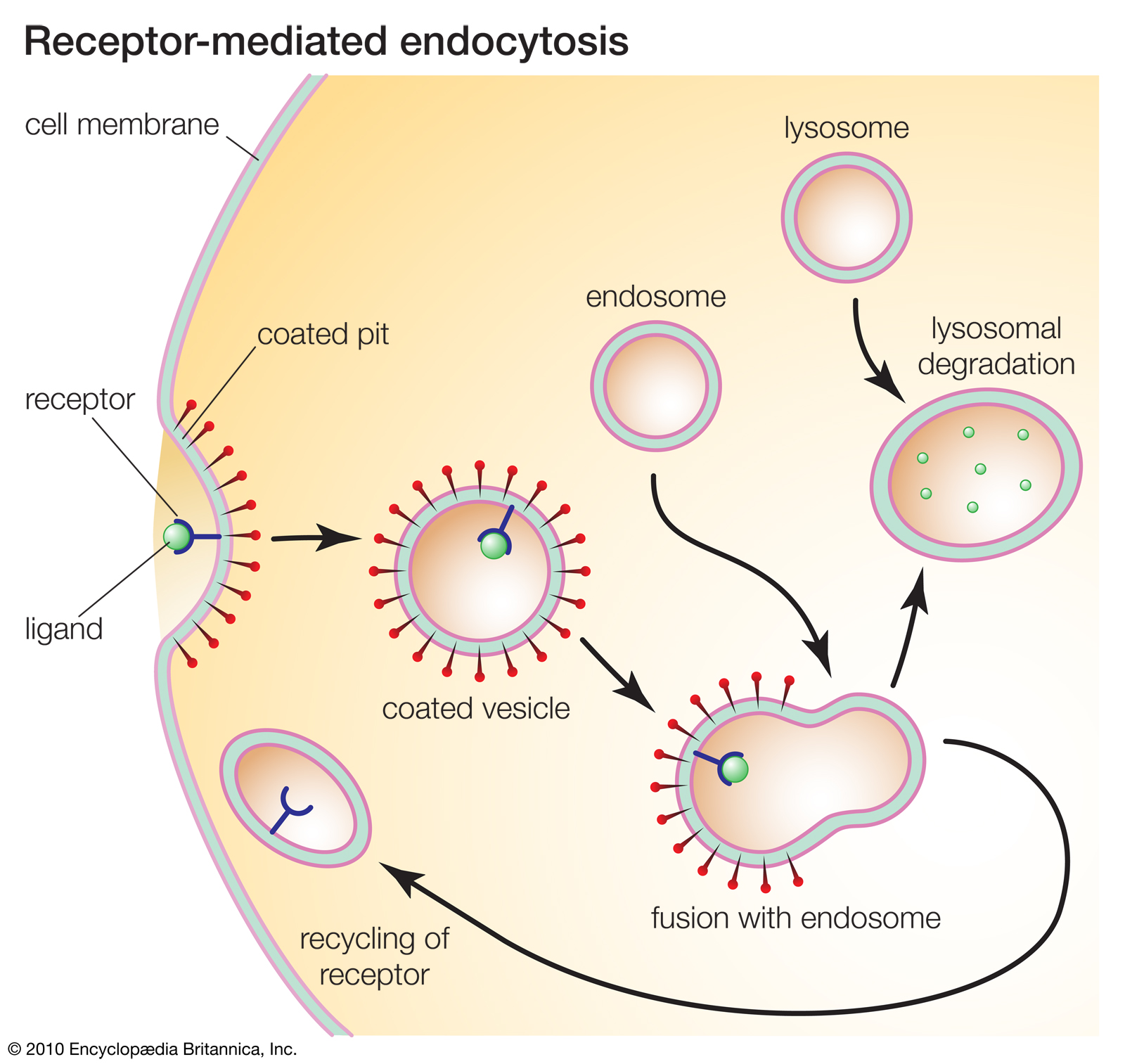
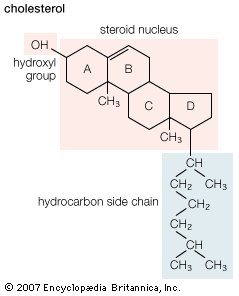
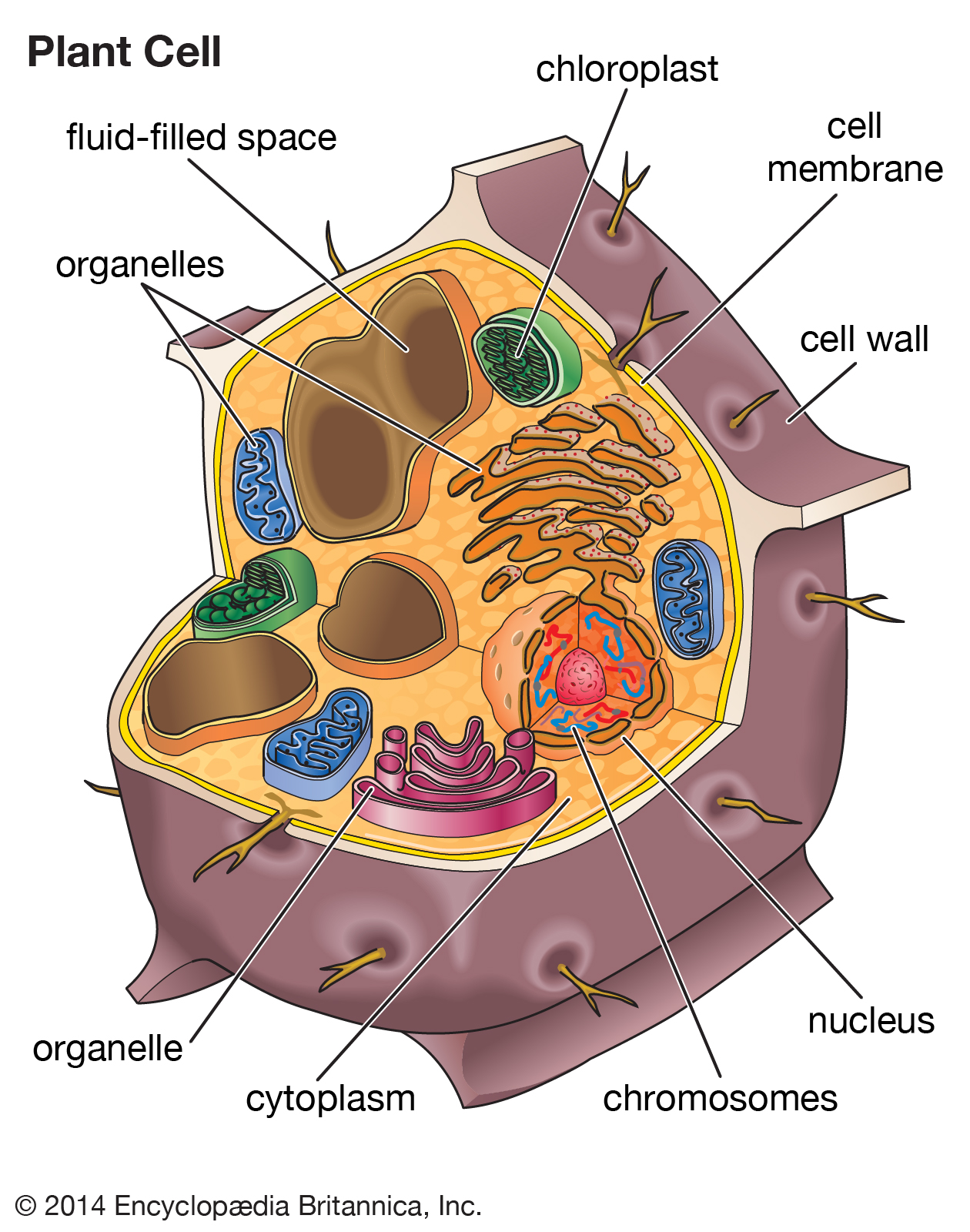
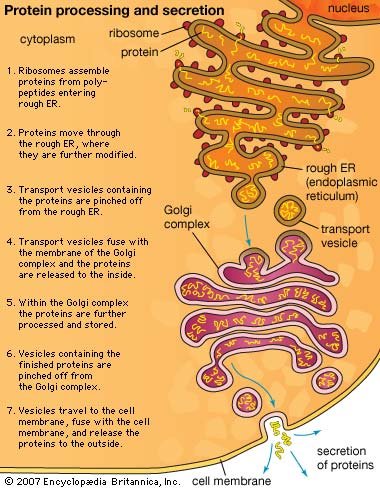
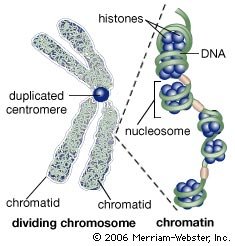
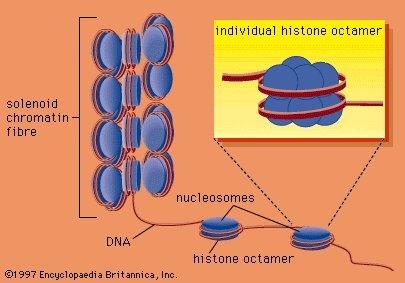
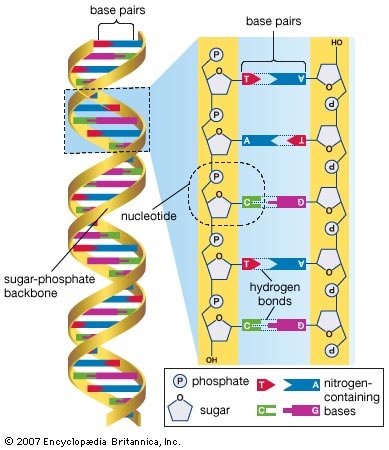
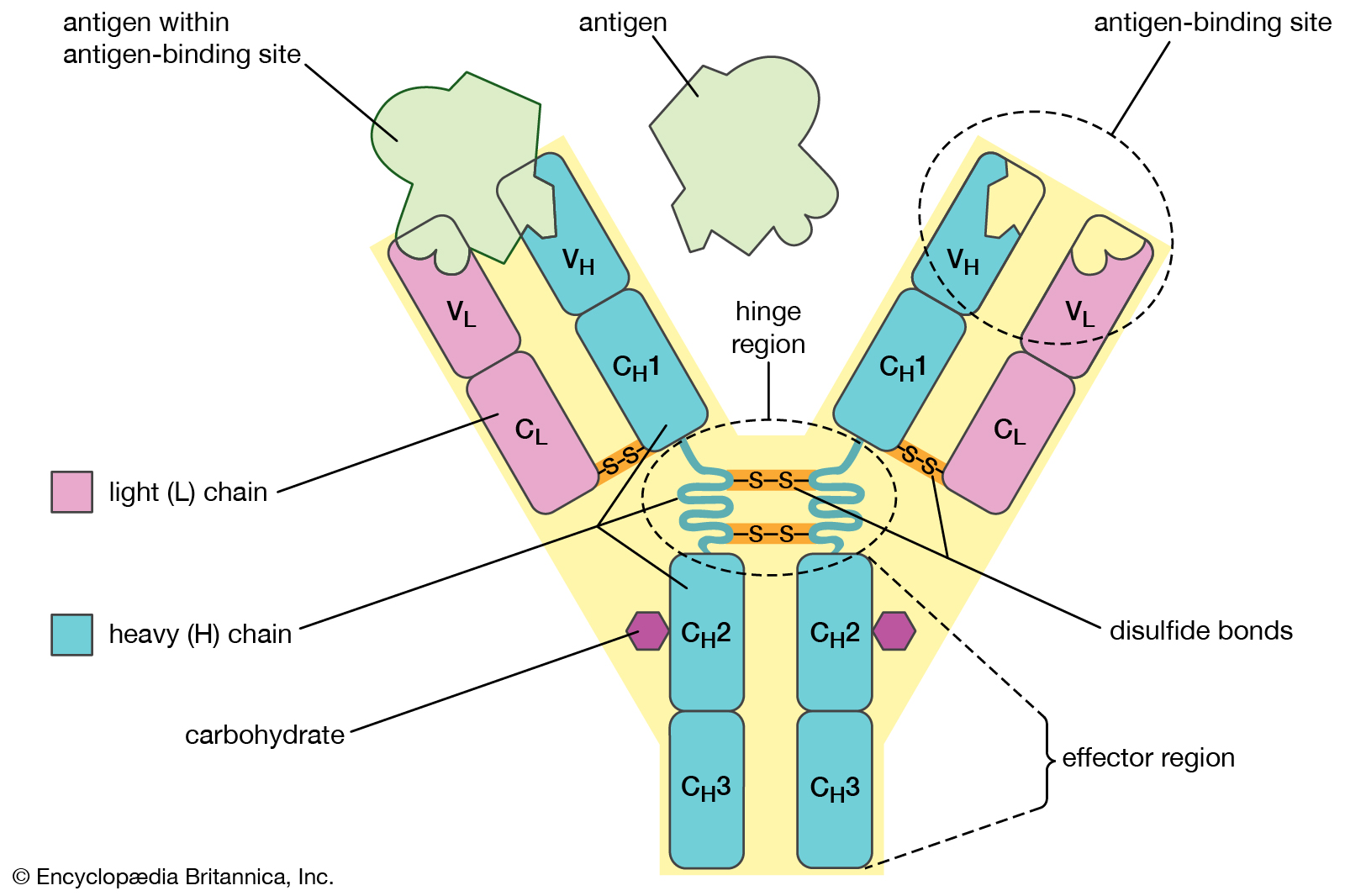
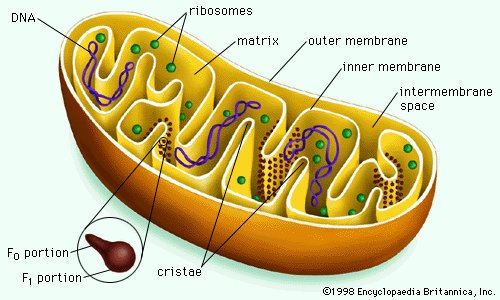
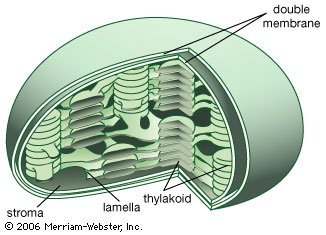
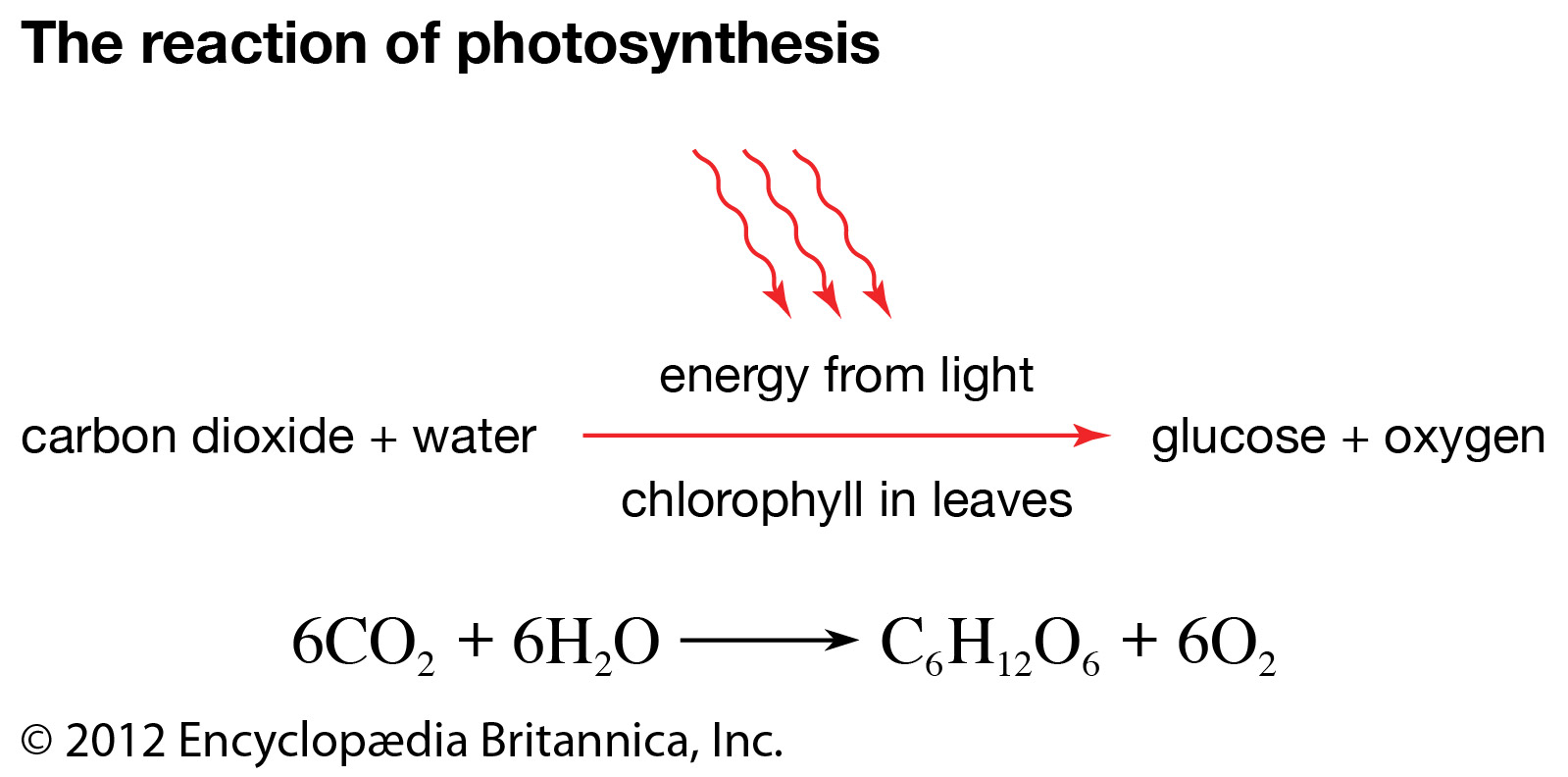

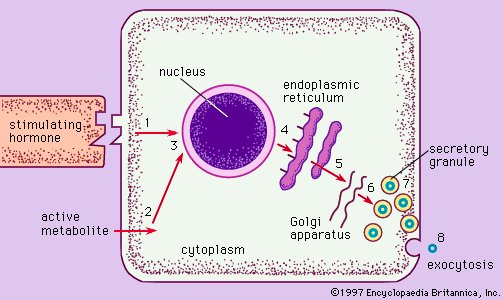
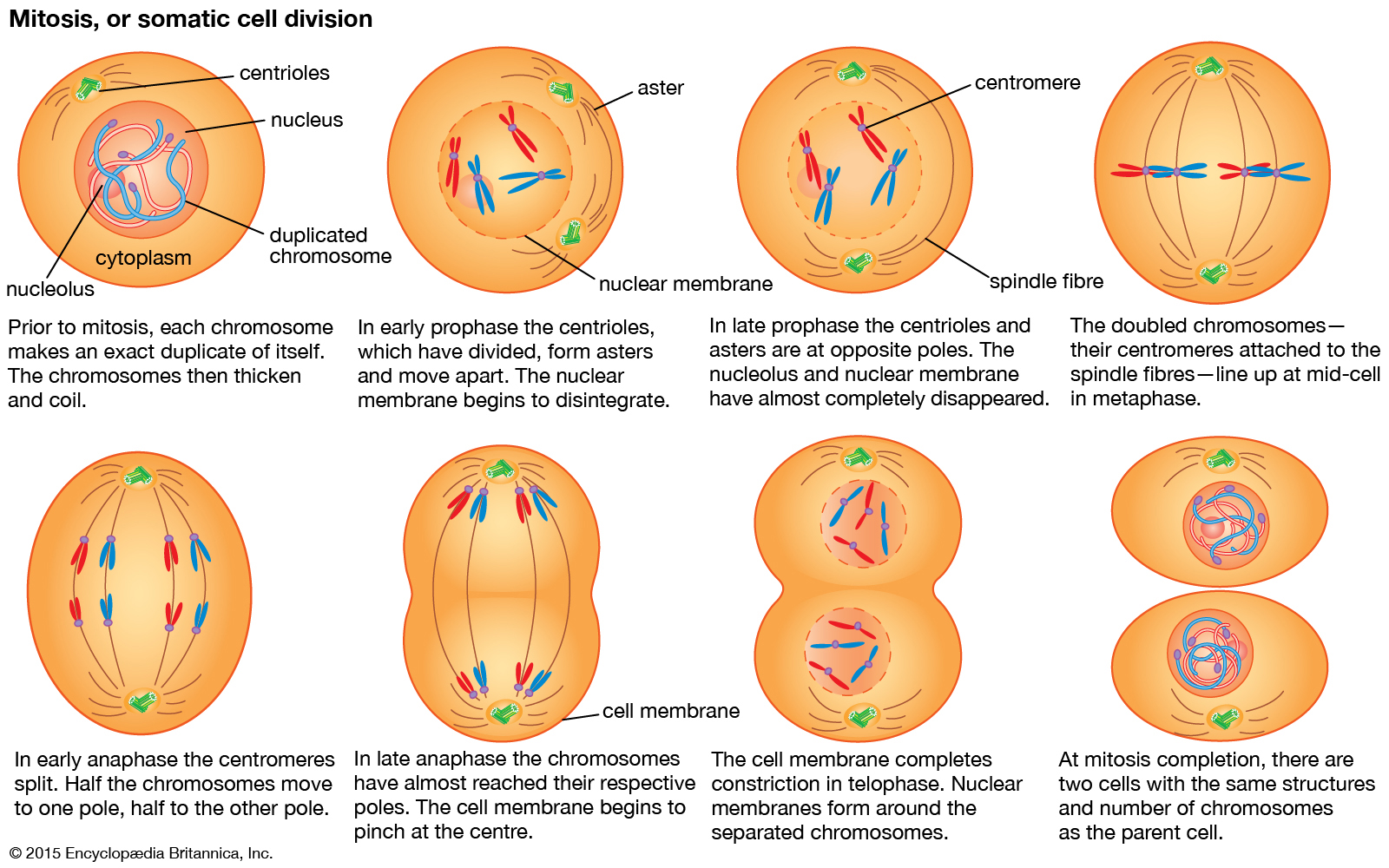
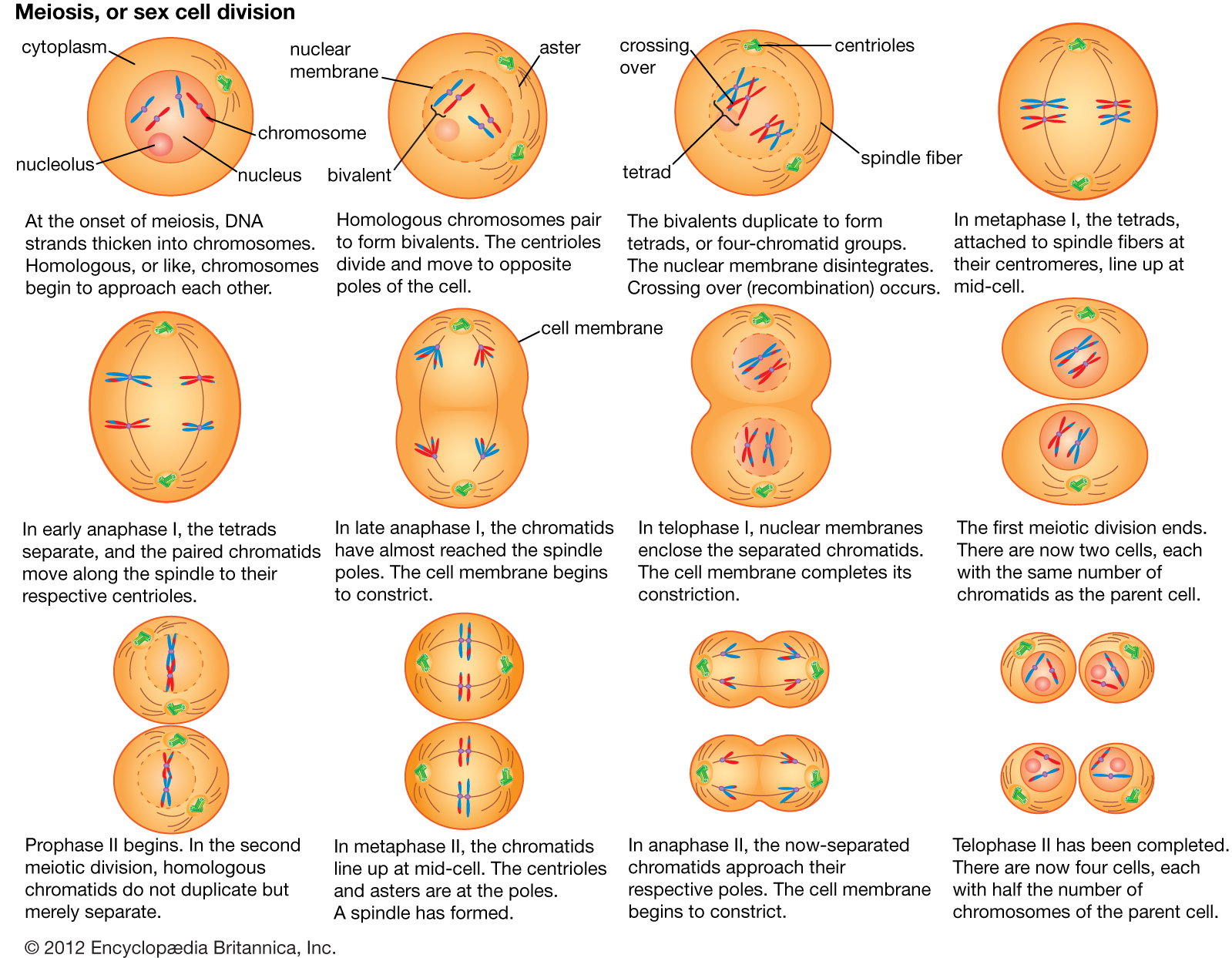
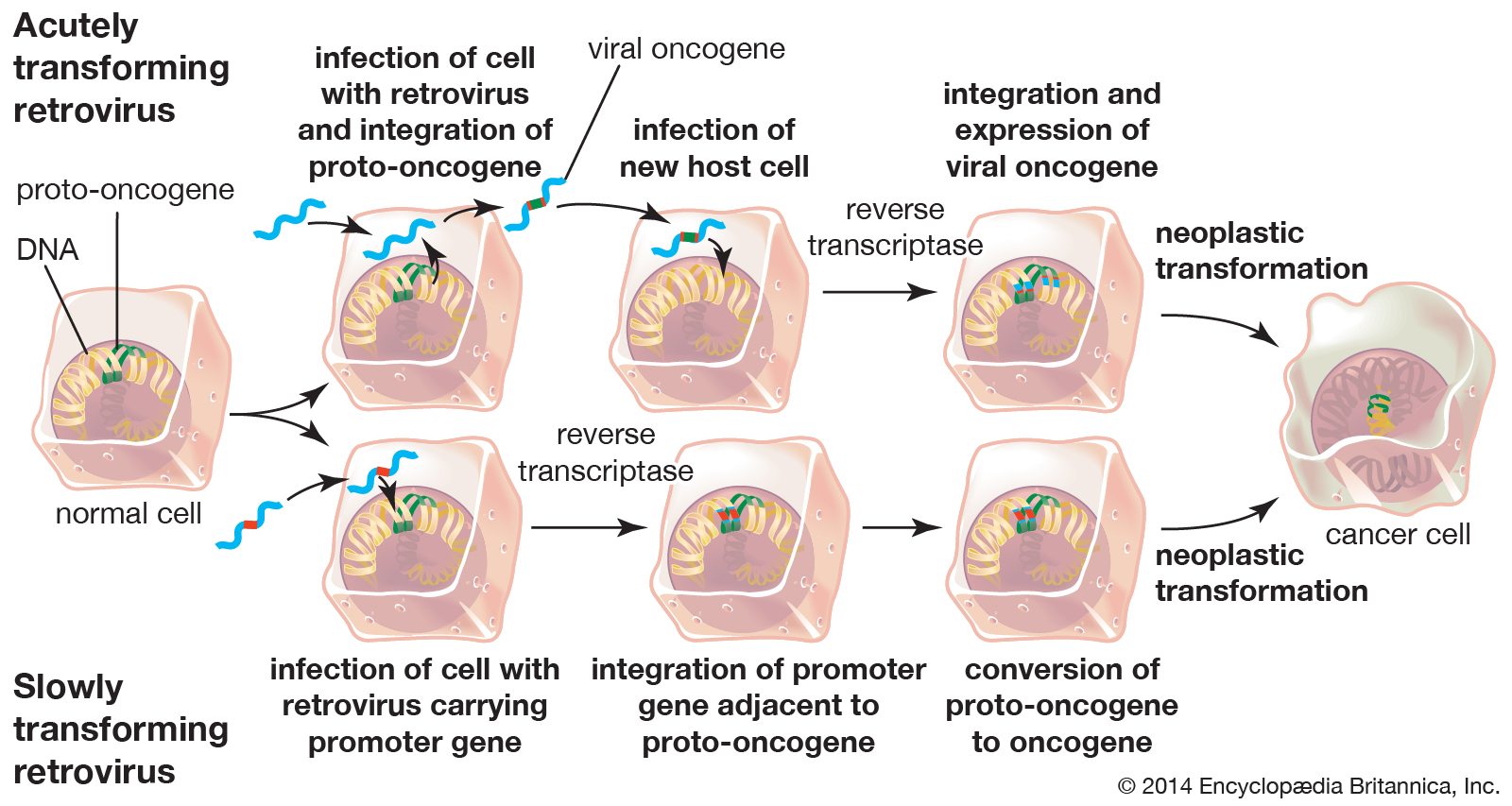
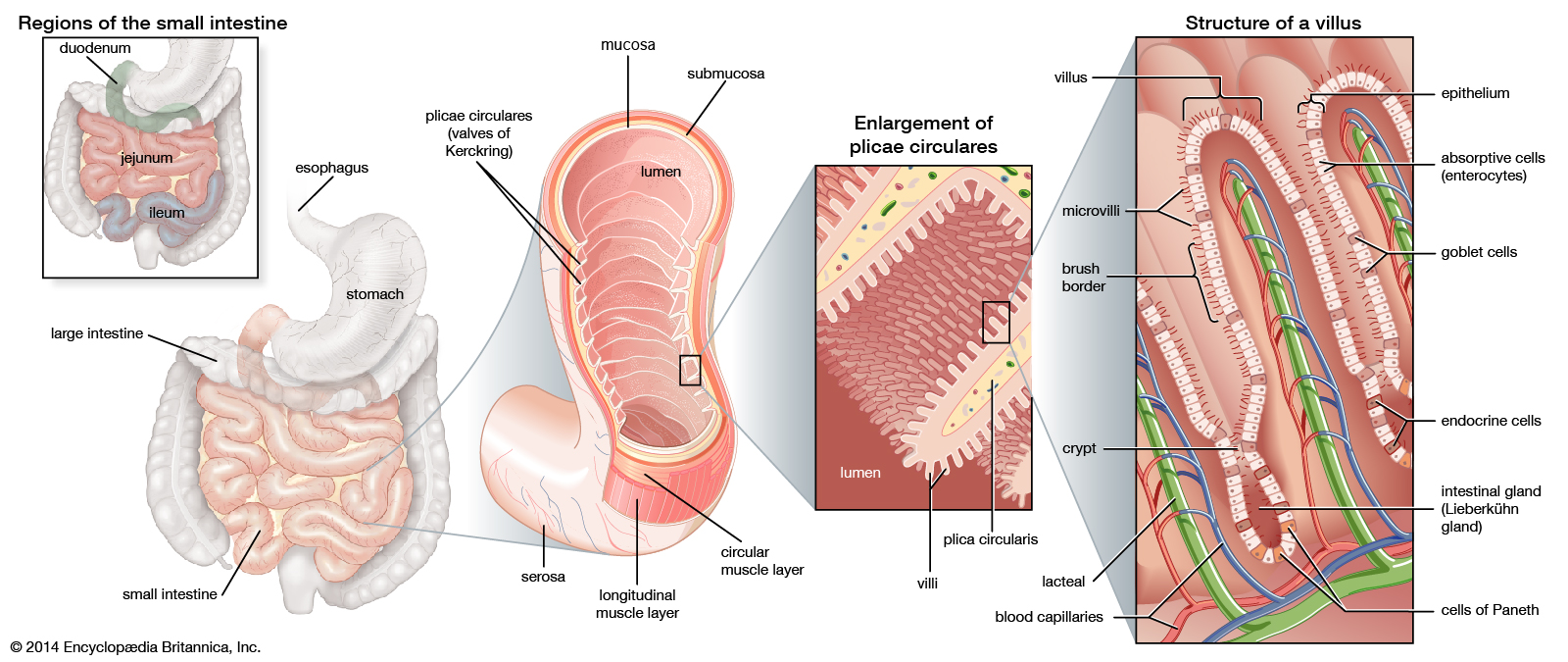
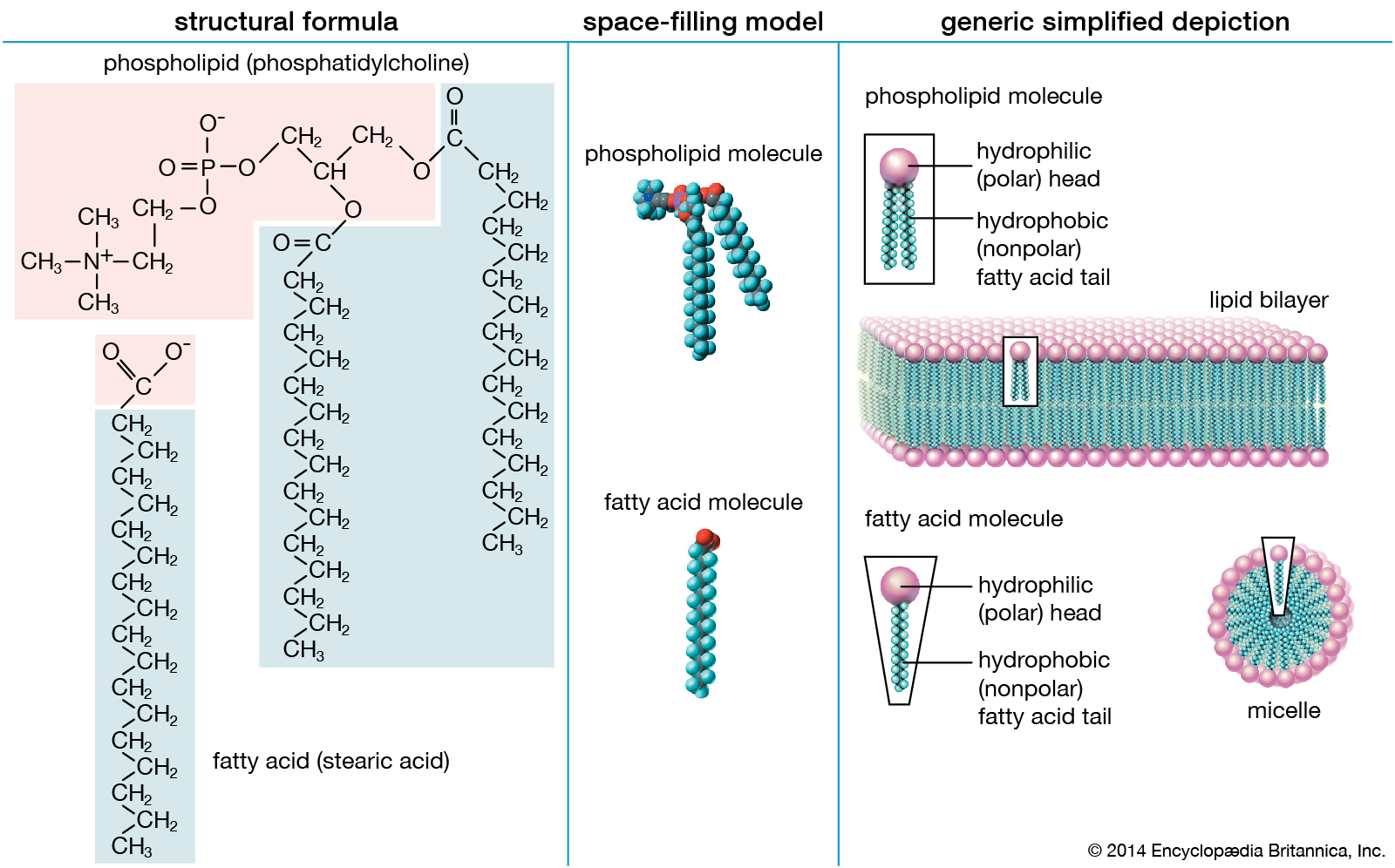





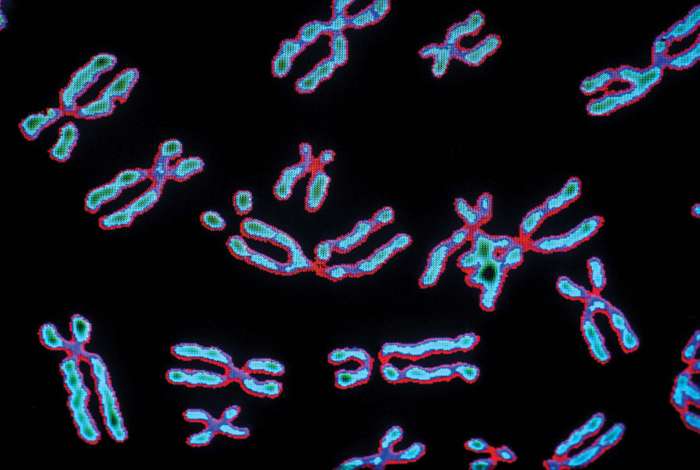


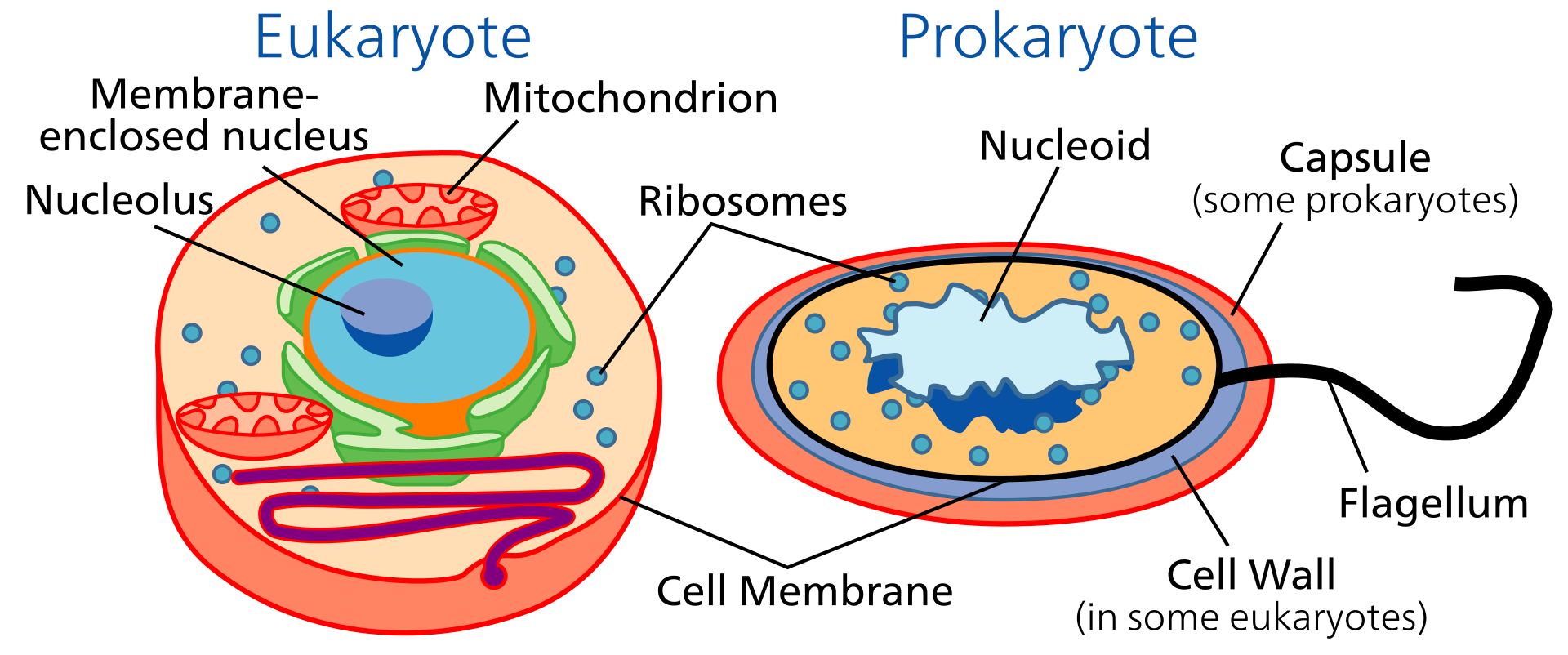
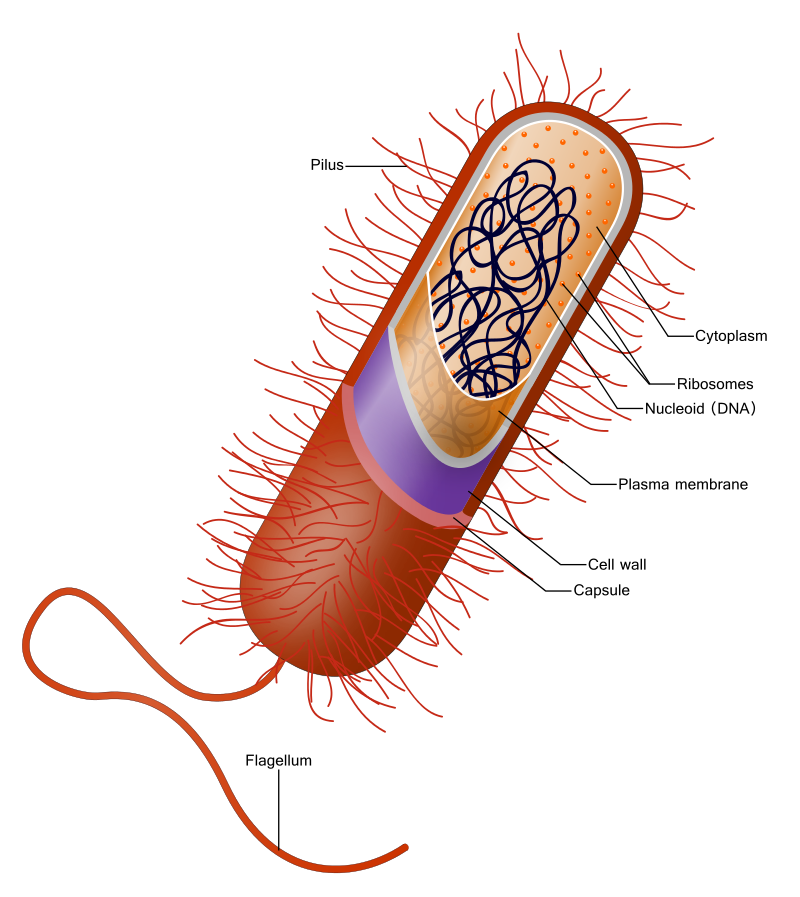
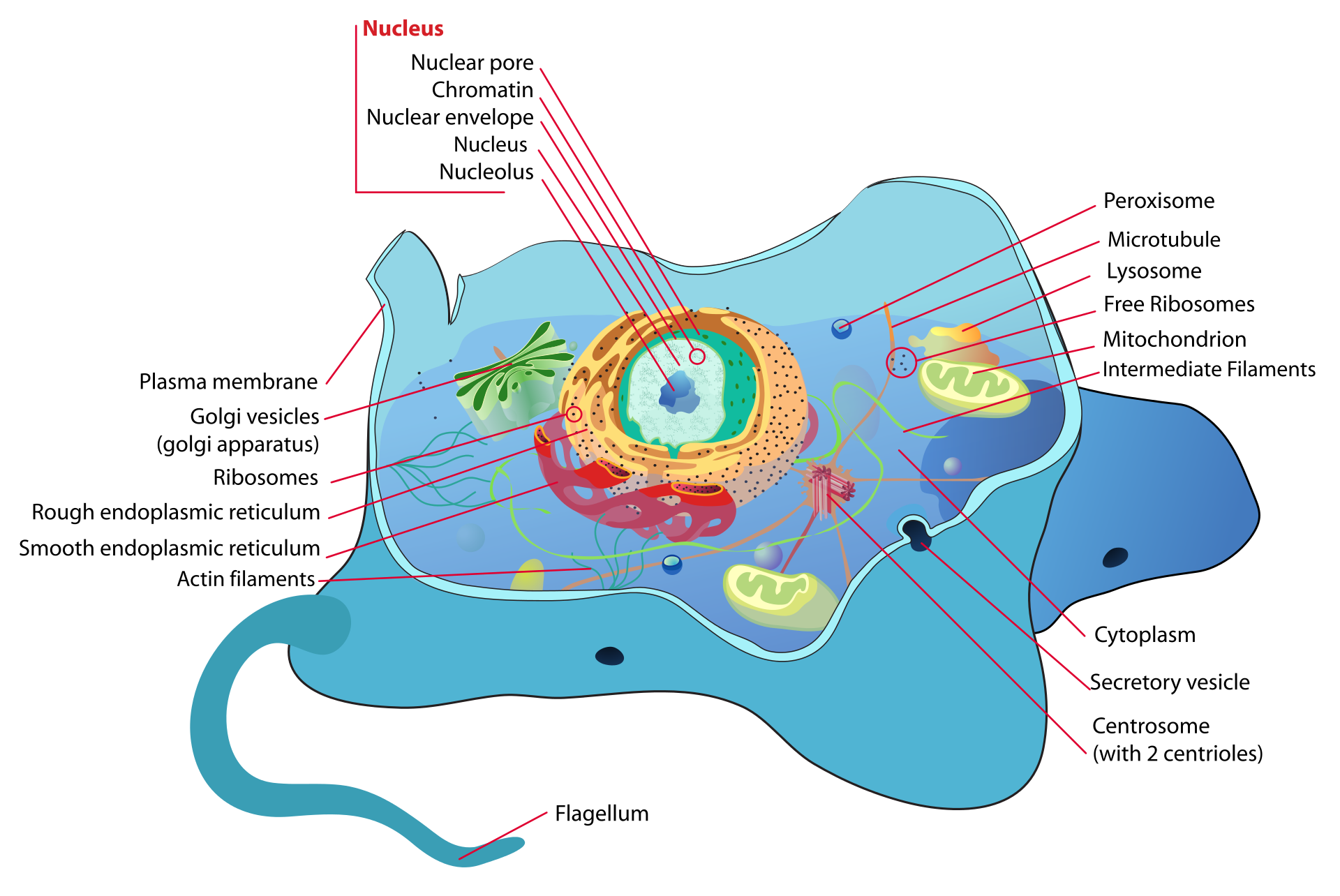
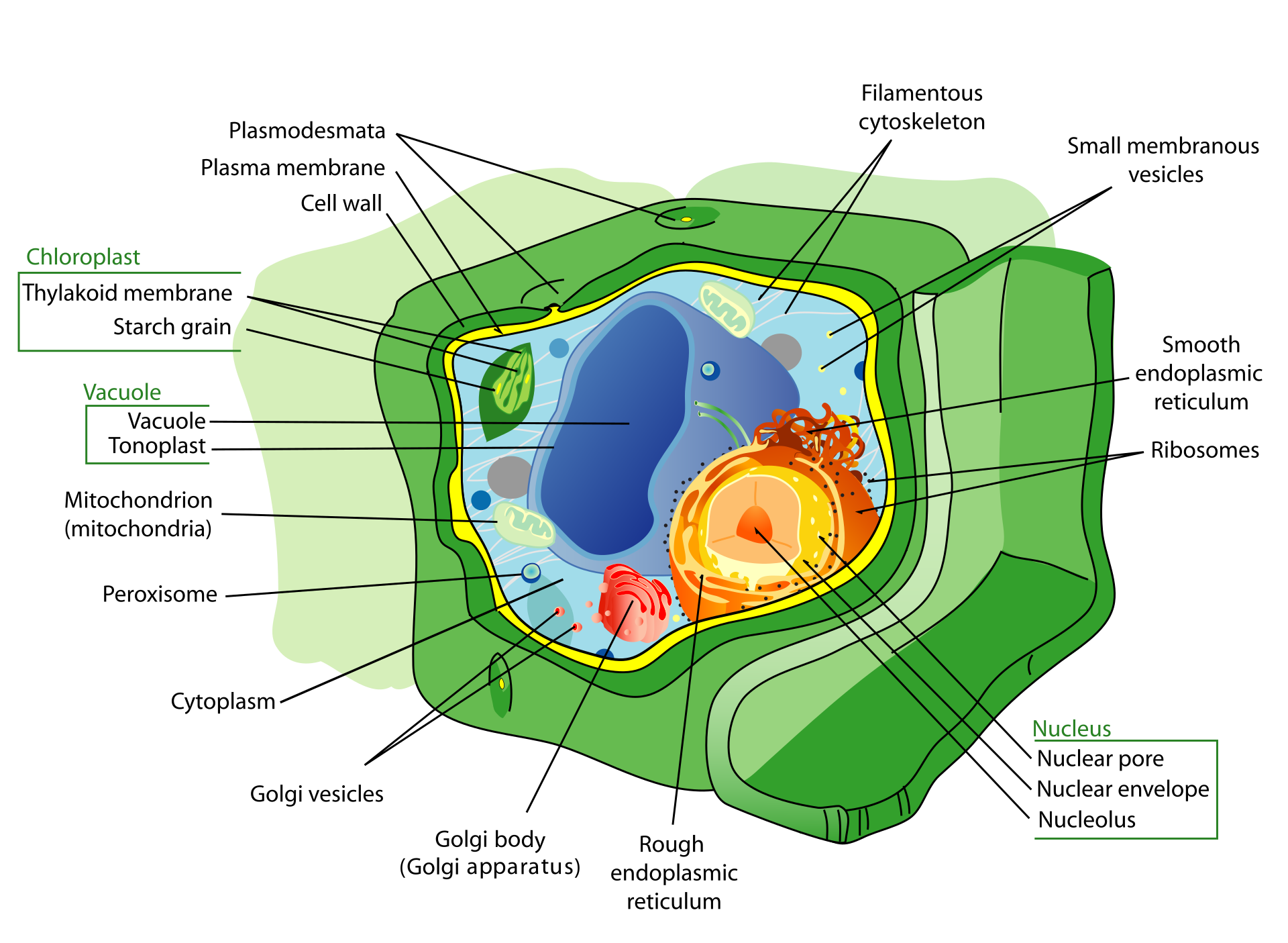
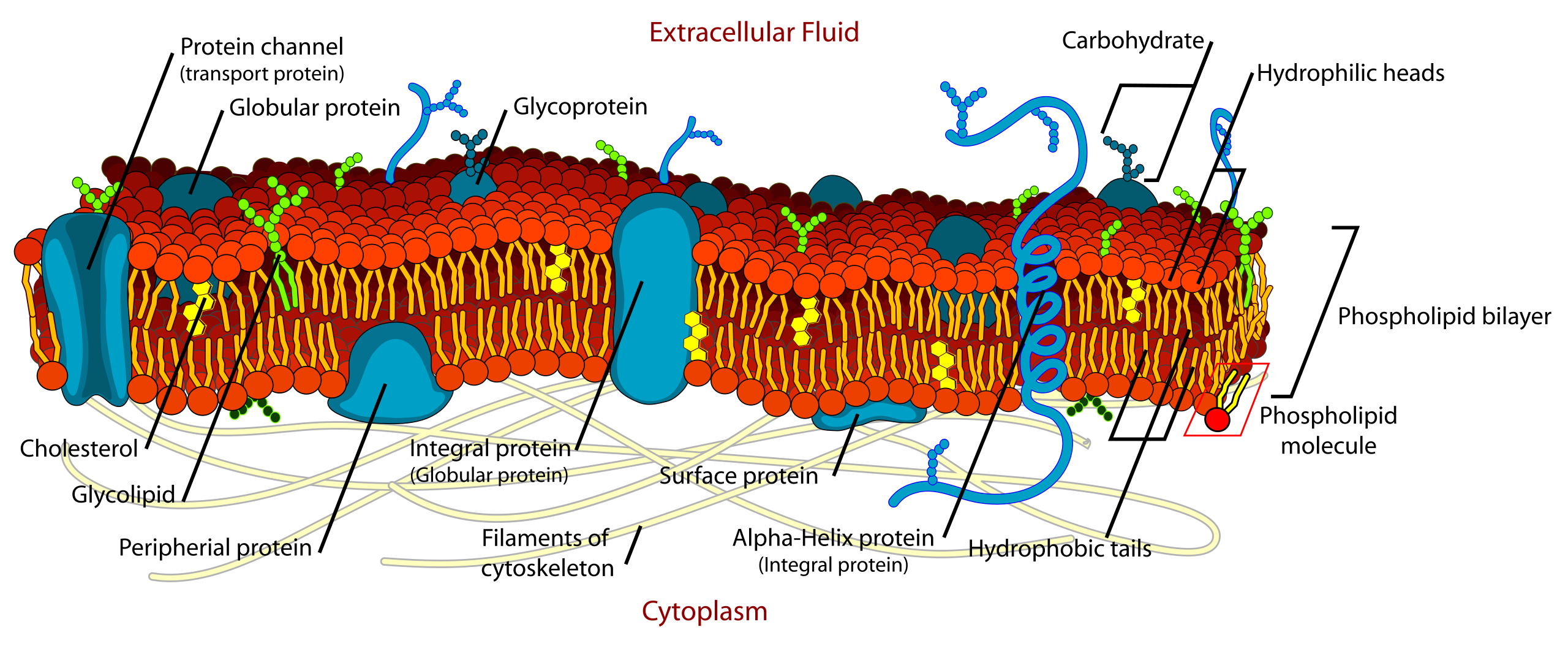
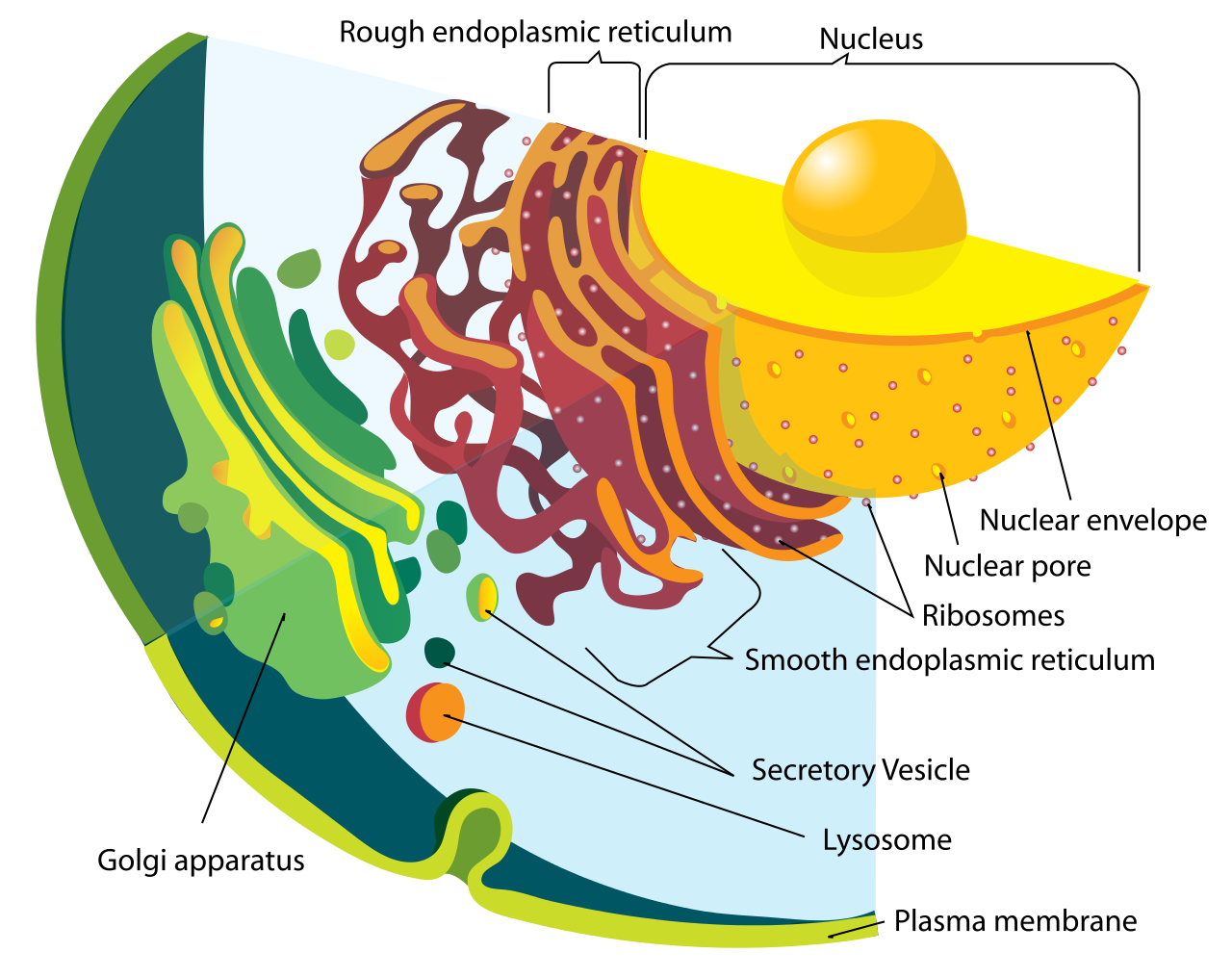
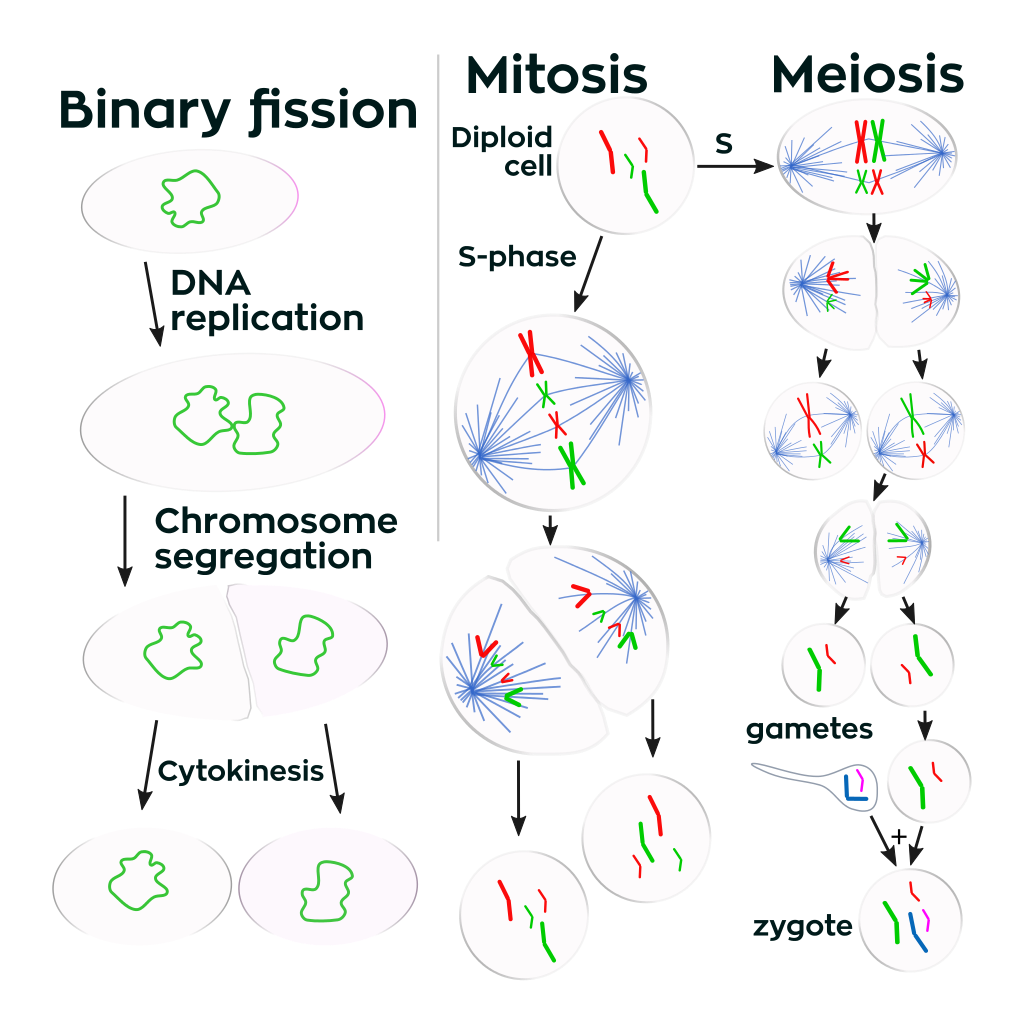
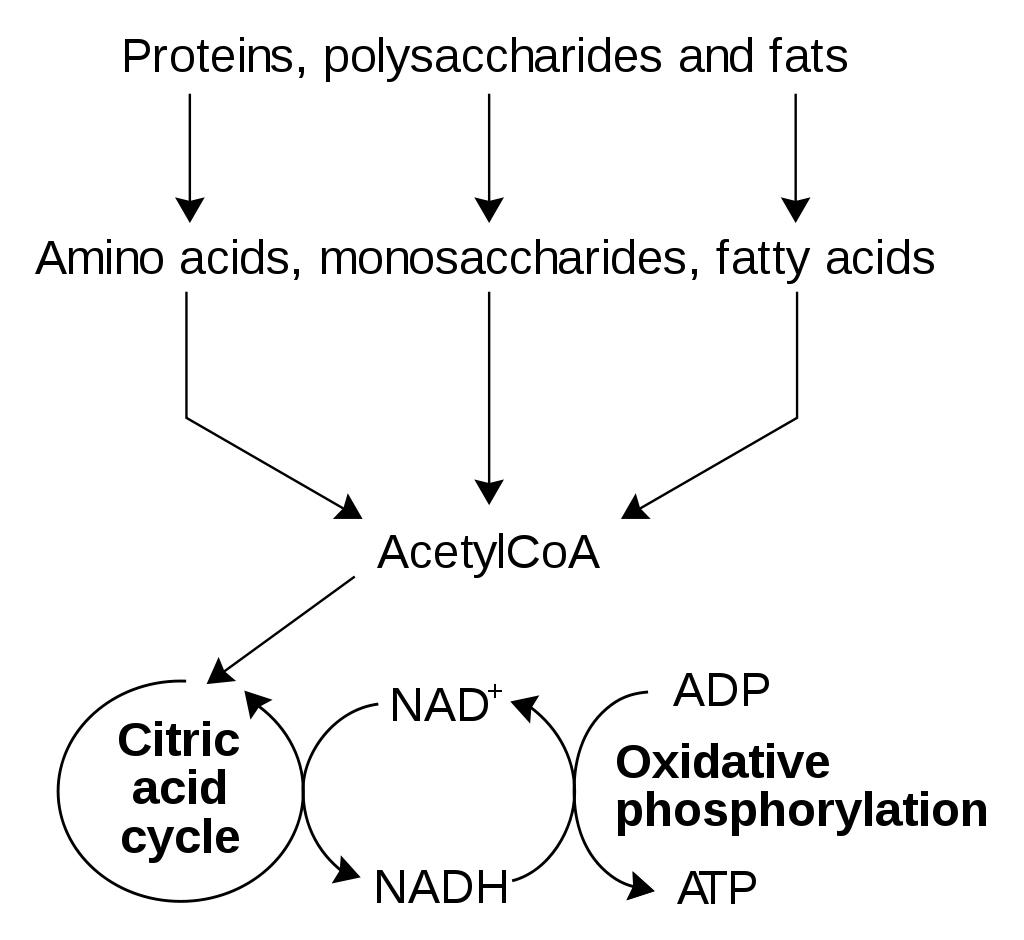
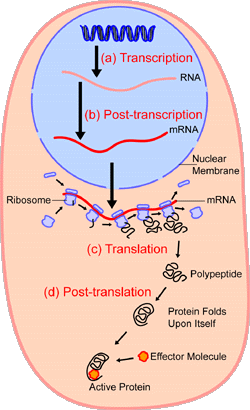

{kind=link}
{kind=link}